, Yves Etienne2 and Maurice Niddam3
(1)
Faculté de Médecine, Marseille, France
(2)
Unité de Médecine Légale, Hôpital de la Timone, Marseille, France
(3)
Unité SAMU 13, Centre 15, Hôpital de la Timone, Marseille, France
There have been few studies of the amygdala’s development in humans to date. However, those that have been carried out are sufficiently exhaustive that we have a fairly accurate idea of this development.
Among excellent former works, the works of Johnston in 1923, Elizabeth Caroline Crosby and Tryphena Humphrey in 1941 and George Macchi in 1951 must be cited. But it is the contemporary authors, notably Fabiola Müller and Ronan O’ Rahilly, who have brought an unrivalled contribution to the field, thanks to modern computer reconstruction processes of serial sectioning performed on significant numbers of embryos. It is since their remarkable work in particular that we have been able to precisely track the development of the amygdala during embryonic development.
3.1 Development During the Embryonic Phase
It is from 5½ weeks into development (stage 16, approx. 6–8 mm) that the primordial, corticomedial and baso-lateral areas of the amygdala develop almost simultaneously (Müller and O’Rahilly 2006).
The first nuclei, however, will not be visible until between stages 17 and 21: recall that at stage 17 (F Müller and R O’Rahilly 1989), the embryo is at the beginning of the 6th week of development and that it measures about 11–14 mm. On a transverse section of the forebrain, through the interventricular foramina, it can be clearly seen that the two lateral ventricles (cavities of the two telencephalic vesicles) communicate widely, despite the presence of the choroid plexus, with the median ventricle (the diencephalic cavity) or third ventricle (Fig. 3.1). The roof of these cerebral ventricles (pallium) is rather thin. It continues laterally and thickens ventrally to form the floor of the telencephalon or subpallium.1 The thickenings of this floor are referred to as the ventricular eminences.2 The side walls of the third ventricle, which follow those of the lateral telencephalic ventricles, are also the thickened zones that will be at the origin of the diencephalic nuclei. It can thus be observed on the telencephalic wall, and projecting into the lateral ventricular cavity, the lateral ventricular eminence appeared at stage 15 (Müller and O’Rahilly 2006), in the diencephalon, the emergent thalamus (the dorsal thalamus and the ventral thalamus separated by the sulcus medius and the intrathalamic zone), the subthalamus and the hypothalamus which project slightly into the cavity of the third ventricle. At the telencephalo-diencephalic junction, a noticeable protrusion, labelled the medial ventricular eminence, appeared from stage 14, the significance of which a number of embryologists have questioned. At this stage, the two lateral and medial ventricular eminences2 remain separated by a sulcus. The medial eminence is particularly important because it is here, as shown by F Müller and R O’ Rahilly, that the first nuclei of the future amygdaloid body appear: the cortical, anterior, baso-lateral and medial nuclei.
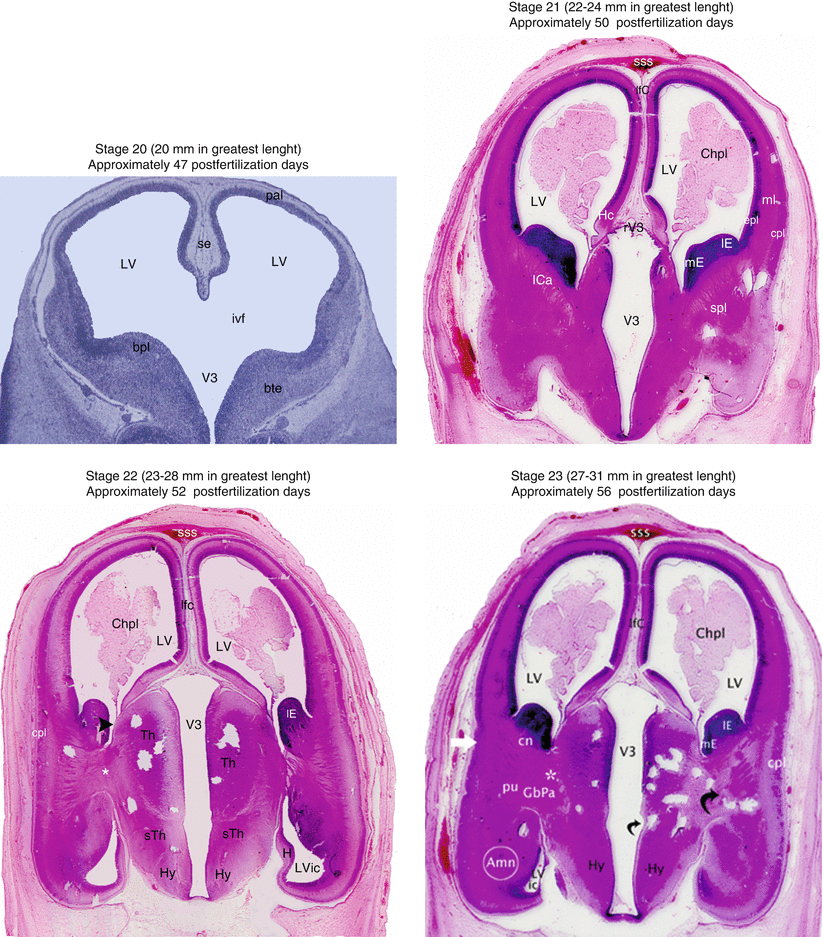
Fig. 3.1
Coronal sections of heads of embryos, showing the evolution of the human brain from stage 20 to stage 23. Amn amygdaloid nuclear complex, bpl basal plate (subpallium), bte basal telencephalon, Chpl choroid plexus, cn caudate nucleus, cpl cortical plate, epl ependymal and mantle layers, GbPa globus pallidus, H hippocampus, Hy hypothalamus, ICa internal capsule, ivf interventricular foramen, lE lateral eminence, lfC longitudinal fissure of cerebrum, LV lateral ventricle, LVic lateral ventricle inferior cornu, mE medial eminence, ml marginal layer, pal pallium, pu putamen, rV3 roof of V3, se septum, spl subpallium, sss superior sagittal sinus, sTh subthalamus, Th thalamus, V 3 third ventricle black arrowhead, sulcus terminalis; little curved black arrow, hypothalamic sulcus; greater curved black arrow, it shows the diencephalon incorporating itself to the telencephalon; white asterisks, they show the lateral migration of the GbPa: white right arrow: future insula
It is the cortical nucleus that appears first, followed by the anterior nucleus, then the baso-lateral and the medial nucleus. The baso-lateral complex is larger than the cortico-medial complex (T Humphrey 1968). The medial nucleus is visible in sagittal sections, in contact with the beginning of the dentate gyrus, already showing the amygdalo-hippocampal proximity!
The medial eminence was previously thought as of telencephalic origin, a notion that remains, still to this day, in a number of books. But the works of F. Muller and R. O Rahilly have indisputably shown since then, thanks to reconstructions, that this eminence is primitively diencephalic even if it tends to invade the telencephalic area early on, becoming di-telencephalic in this manner.3 The amygdala arising from this medial eminence is therefore originally a nuclear diencephalic complex.4 The line of demarcation that separates the telencephalon and the diencephalon is a straight line that links the velum transversum to the preoptic recess: that which lies above this border belongs to the telencephalon. All that is underneath is diencephalic. The medial eminence, by its own expansion, will be quickly divided in two by this line, which explains the discussion on the telencephalic or diencephalic origin of the amygdala. The lateral eminence, originally telencephalic, is smaller. It remains in the subpallium and bulges slightly into the lateral ventricular cavity. As soon as the ventricular eminences cross, the interventricular foramina contract, a phenomenon further accentuated by the expansion of the choroid plexus.
At stage 18 (about 13–17 mm), that is, at 6½ weeks of development, the four nuclei of the amygdala are still inside the medial eminence, but the cortical nucleus extends toward the lateral eminence which therefore contributes partially to its formation. This stage 18 marks the start of the rapid development of the hemispheres, which will be further accentuated from stage 20. Muller and O’Rahilly (1990a) have observed at stage 18 the presence of nervous fibres linking the lateral olfactory stria and the olfactory tubercle at the medial amygdala nucleus, as well as linking the olfactory bulb with baso-lateral and anterior nuclei. Other connections are made with the ventral thalamus via the lateral forebrain bundle.
Stage 19 (about 16–18 mm), that is, between 6½ and 7 weeks of development, could be considered to be the stage of striae. Indeed, it is at this stage that the fibres of the stria terminalis appear. It is also the stage that other fibres of the stria linking the baso-lateral nucleus of the amygdala to the lateral nucleus of the habenula take the path of the medullary stria of the thalamus. The lateral eminence crosses rostrally and from there pushes the medial eminence towards the median line, causing it to protrude slightly into the ventricle. The lateral globus pallidus appears inside the diencephalon, in the subthalamic region (Muller and O’ Rahilly, 1990a). It is also of note, according to these authors, that the nucleus accumbens is recognisable at this stage.
At stage 20 (about 18–22 mm), the 7th week of development, the medial eminence is entirely in the telencephalic territory. The nuclear complex of the amygdala is from now positioned in the middle part of the subpallium. Other nuclei have appeared: those of the diagonal band and of the septum. Connections are established between the two latter structures, the amygdala nuclei, the hippocampus, the diencephalic structures and the bundle of the prosencephalon (Muller and O’Rahilly 1990a).
At stage 21 (about 22–24 mm), towards the 50th day of development, the future medial globus pallidus starts to differentiate, in the diencephalon. The bundle of the prosencephalon that unites the telencephalon and the diencephalon is at this stage highly visible (Muller and O’ Rahilly 1990b).
Stage 22 (about 23–38 mm), towards the 52nd day of development, the hemispheric neopallium is well developed and the internal capsule that leaves it, is visible. The lateral globus pallidus begins its migration towards the telencephalon.
Stage 23 (about 27–31 mm), towards the 56th day, corresponds to the end of the embryonic period. The putamen that arises from the subventricular layer of the lateral eminence is visible from this stage, and due to its rapidly increasing volume, it requires the amygdala, within which the significant central nucleus is formed, to move towards the temporal area. The caudate nucleus itself also becomes morphologically recognisable. The lateral globus pallidus migrates towards the putamen. It is also the stage at which the insula differentiates, beginning to form the lateral or sylvian fossa (compared to the claustrum, which had appeared at stage 22), that the emergent dorsal and ventral thalamic structures appear, that the rhombencephalic nuclei organise and that the anterior commissure is formed (O’Rahilly and Müller 1999). It is at the final stage in which the hippocampus positions itself in proximity to the future temporal pole.
Related posts:
 From Prefrontal Lobectomies to Amygdalectomies
Anatomy of Emotions via the History of Neuroanatomy and Neurosciences
From Prefrontal Lobectomies to Amygdalectomies
Anatomy of Emotions via the History of Neuroanatomy and Neurosciences
 Vascularization of the Cerebral Amygdala
Vascularization of the Cerebral Amygdala
 The Bed Nucleus of the Stria Terminalis
The Bed Nucleus of the Stria Terminalis
 Technique for Dissecting the Amygdaloid Body and Its Close Connections
Technique for Dissecting the Amygdaloid Body and Its Close Connections
 The Concept of “Extended Amygdala”
The Concept of “Extended Amygdala”
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


