, Yves Etienne2 and Maurice Niddam3
(1)
Faculté de Médecine, Marseille, France
(2)
Unité de Médecine Légale, Hôpital de la Timone, Marseille, France
(3)
Unité SAMU 13, Centre 15, Hôpital de la Timone, Marseille, France
10.1 Generalities
These terms and their abbreviation in English “NBST or BST” are used to refer to a practically seamless continuum of neuronal cell clusters bordering the concave edge of the caudate nucleus, from its head to its tail, the septal region of the amygdaloid body, and within which passes the stria terminalis.
Various names have been used to refer to these cell clusters such as the “support nucleus” (M Volsch 1910) or “secondary nuclei of the stria” (CWA Kappers 1921) or even the “enveloping nucleus” (EW Lauer 1945) and “interstitial nuclei” (R Diepen 1962).
In this bed, in addition to the stria terminalis (see Chap. 6), the large thalamo-striate vein and its tributaries are routed before it drains into the internal cerebral vein. Cell clusters of the BNST are in contact with the two aforementioned elements and are usually interposed there between.
The BST, which we seem to have more or less rediscovered in recent years, is already well known from the work mentioned above and especially since the memorable anatomy works performed by JB Johnston, in 1923. In a long article devoted to the evolution of the forebrain, the author described this structure from the early stages of embryonic life and showed its central position and its role: establishing communication between the amygdaloid complex and the olfactory area. Subsequently, many contributors have clarified the anatomy of the BST, including OJ Andy and H Stephan, in 1968, who studied its paraseptal portion; F Sanides, in 1957, who studied its caudal portion; and, above all, H Strenge et al., in 1977, who carried out an exhaustive and accurate histological study of the entire cell mass using serial sections perpendicular to the arch that forms the BST. We should also recall the work of JS De Olmos (2004) who studied the connections between the BST and the amygdala in particular.
Three portions are usually identified for the BST: the rostro-medial portion, the intermediate portion and the caudal portion.
The rostro-medial portion is special. It forms three areas: the most anterior area, sometimes referred to as “pre-septal”; the second area located behind the previous area, often referred to as the “commissural” considering its proximity to the anterior commissure; and the third area, the “retro-commissural” area.
At the level of pre-septal area, the BST is in the centre of the striatum and positioned medially with respect to the anterior branch of the internal capsule. It insinuates itself between the caudate nucleus, element of the dorsal striatum and the nucleus accumbens (Fig. 10.1a, a’), element of the ventral striatum which has just received one or more branches of the stria terminalis. It is medially covered by subependymal epithelium, which forms, at this level, the inferior recess of the front horn (or anterior cornu) of the lateral ventricle.
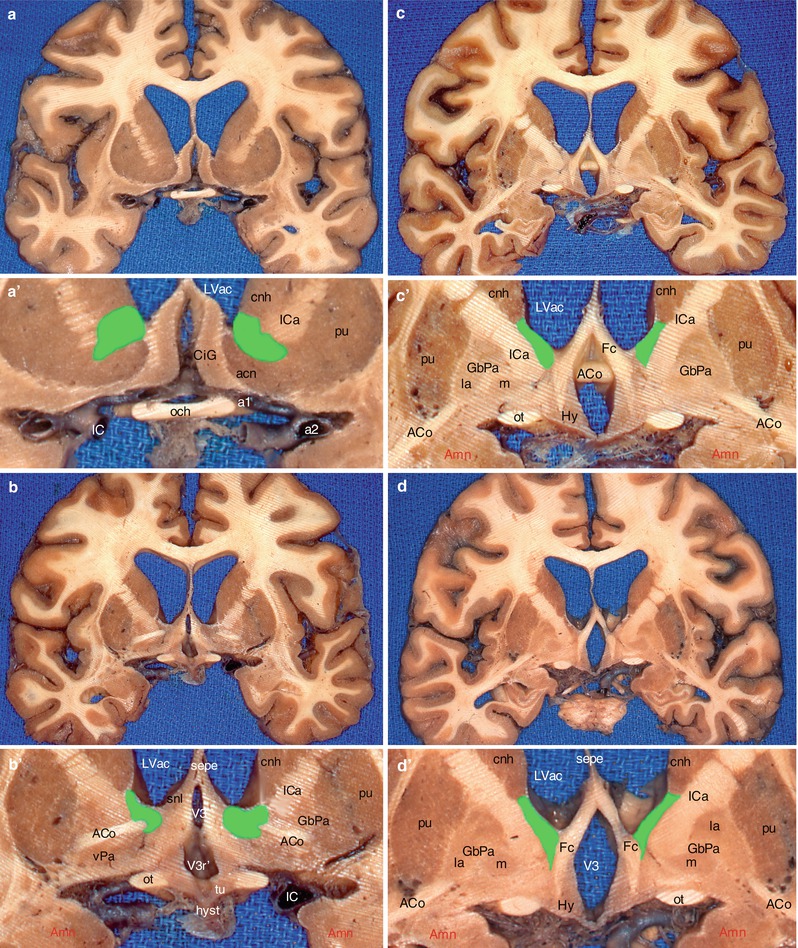
Fig. 10.1
Location of the BST on a series of brain’s coronal sections in the area of the anterior commissure. (a) Section through the optic chiasm; (b) section through the optic recess of V3; (c) section behind the median part of ACo; (d) section through the hippocampal digitations; (a′–d′) enlargements of a, b, c and d, showing the green-coloured BST. acn accumbens nucleus, ACo anterior commissure, Amn amygdaloid nuclear complex, CiG cingulate gyrus, cnh caudate nucleus head, Fc column of fornix, GbPa globus pallidus, la pars lateralis, m pars medialis, Hy hypothalamus, hyst hypophyseal stalk, IC internal carotid, ICa internal capsule, LVac lateral ventricle, anterior cornu, och optic chiasm, ot optic tract, pu putamen, sepe septum pellucidum, snl septal nucleus (lateralis), tu tuber, V3 third ventricle, V3 r′ optic recess, vPa ventral pallidum, a1 anterior cerebral artery, a2 middle cerebral artery
At the commissural level, the BST always underlies the caudate nucleus and is positioned against the lateral ventricular wall, inside the crus anterior of the internal capsule. It remains in a dorsal position with respect to the anterior commissure, which forms its median trajectory, slightly arched, with a dorsal convexity (Fig. 10.1b, b’ and c, c’ and Fig.10.2).
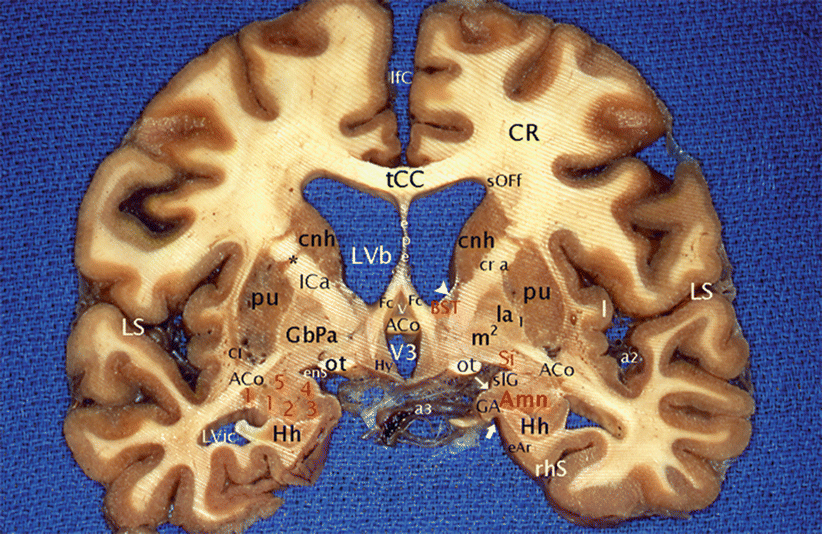
Fig. 10.2
The anterior commissure, the amygdalae and the BST on a coronal section of the brain through the columns of the fornix. ACo anterior commissure, Amn (red keys) amygdaloid nuclear complex; 1 lateral, 2 baso-lateral, 3 baso-medial, 4 cortical, 5 central; BST (red key) bed nucleus of the stria terminalis, Cl claustrum, cnh caudatus nucleus (head), CR corona radiata, eAr entorhinal area, enS endorhinal sulcus, Fc column of the fornix, GA gyrus ambiens, GbPa globus pallidus, la pars lateralis, m pars medialis, 1 external medullary lamina, 2 internal medullary lamina, Hh hippocampus head, Hy hypothalamus, I insula, ICa internal capsule (cr a crus anterior), lfC longitudinal fissure of the cerebrum, LS lateral sulcus, LVb lateral ventricle body, LVic lateral ventricle, inferior cornu; ot optic tract, pu putamen, rhS rhinal sulcus, sepe septum pellucidum, Si (red key) substantia innominata, slG semi-lunaris gyrus, sOFf superior occipito-frontal fasciculus, tCC truncus of the corpus callosum, v vulva, V3 third ventricle, a2 middle cerebral artery, a3 posterior cerebral artery, *putamino-caudate bridge, thin white arrow semi-annular sulcus, thick white arrow uncal notch, white arrowhead: la lamina affixa
At the rear of the anterior commissure, the BST remains connected to the head of the caudate nucleus, which it follows up to the posterior third (Fig. 10.1d, d’).
For R Walter et al. (1991), who have studied the differential distribution of histochemical markers in the BST of the human brain, this rostro-medial portion is the part which specifically corresponds to the BST due to the fact that beyond this point, the presence of the dorsal part of the magno-prothalamic and parvocellular nuclei, initially described by H Brockaus, makes it difficult to interpret the results. Thus, the marks of A Walter et al. only concern the rostro-medial part of the NBST.
The intermediate portion (see Fig. 7.15a) is the longest of the three portions of the BST. It follows the entire body of the caudate nucleus, i.e. almost half of the caudate arch. The front concave trajectory includes three segments, the superior segment with an inferior concavity, the middle segment with an anterior concavity and the inferior segment with a downward, almost straight trajectory.
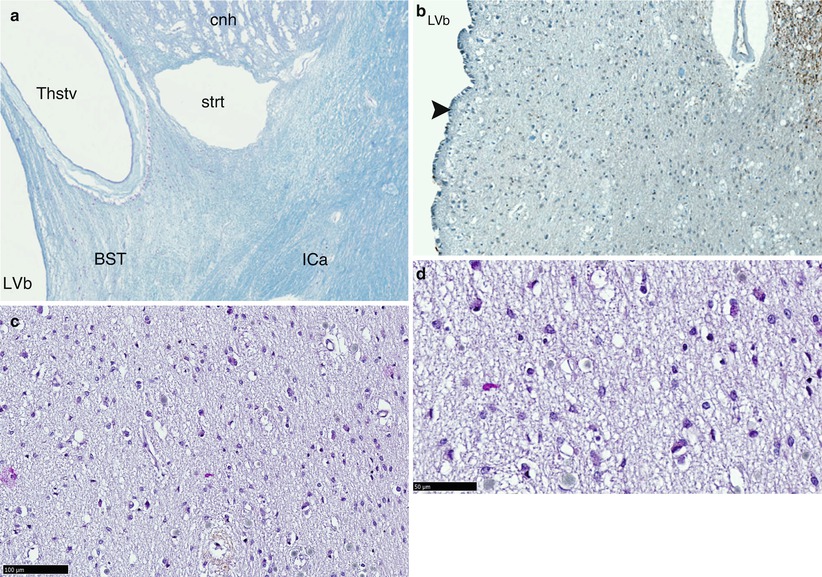
This section begins beyond the head of the nucleus where the thalamus contacts the body of the caudate nucleus. A thalamo-caudate sinus forms immediately and extends along a groove in a circular arch to the tail of the caudate nucleus. The cells, which form the BST and especially those adjacent to the thalamus, play an important role such as demonstrated by numerous recent studies. This is not surprising if we refer to embryology as it is clear that the thalamo-caudate sinus is a strategic area where the striatum and thalamus become closer to each other so as to allow the diencephalo-telencephalic junction and the incorporation of the diencephalon in the median brain (see Chap. 3). At the level of the dorsal surface of the thalamus, the furrow occupied by the BST is also bordered, on the ventricular side, by a thin white substance lamina, the lamina affixa or lamina cornea, an embryonic residue of the telencephalic white substance separating the striated body from the thalamus. The role played by this lamina affixa is to attach the thalamo-striate vein to the bottom of the furrow. It also transforms this furrow into a triangle, which corresponds to the location of the BST. This triangle will also expand into a square or rectangle according to the areas being observed (Fig. 10.3
).
Fig. 10.3
Microscopic aspects of the BST (bed nucleus of the stria terminalis) in front of the anterior commissure. (a) Location of BST on a coronal section, (b) low-power view (X100), (c) intermediate-power view (×200), (d) high-power view (×400): we observe little scattered neurons. BST bed nucleus of the stria terminalis, cnh caudate nucleus head, ICa internal capsule, LVb lateral ventricle body, strt stria terminalis, Thstv thalamo-striate vein, black arrowhead ependymal layer
Beyond this point and especially at the level of the posterior surface of the thalamus, the lamina affixa disappears and the ventricular wall of the BST consists only of the ependymal layer.
The inferior portion, although close to the amygdala, is smaller in size. Its volume seems to decrease proportionally to the volumetric reduction of the caudate nucleus which tapers progressively towards its tail section.
The NBST cells (Figs. 10.3 and 10.4) have been described by H Strenge et al. (1977):
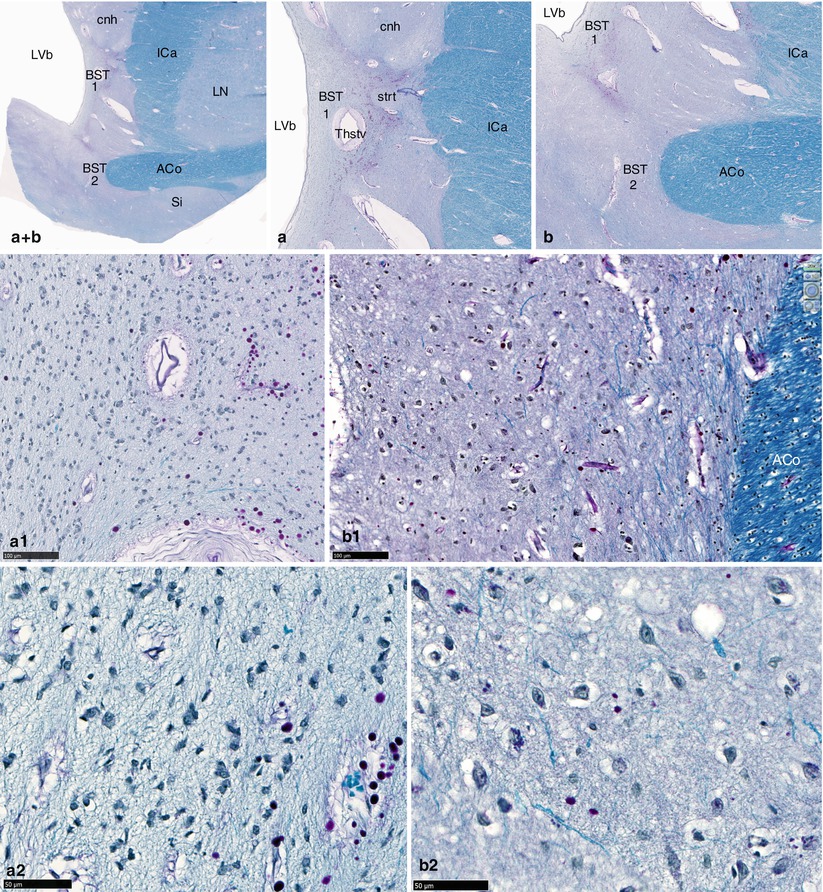
Fig. 10.4
Microscopic aspects of the BST (bed nucleus of the stria terminalis) through the anterior commissure. (a + b) Global view of the BST; (a) BST under the head of the caudate nucleus (BST1); (b) BST around the medial end of the left part of ACo (BST2); (a1, a2, b1, b2) luxol fast blue–PAS stainings (×200, ×400, ×200, ×400), of BST 1 (a1, a2) and BST 2 (b1, b2). ACo anterior commissure, BST bed nucleus of the stria terminalis, BST1 pars under caudate nucleus, BST2 pars around the ACo, cnh caudate nucleus head, ICa internal capsule, LN lentiform nucleus, LVb lateral ventricle body, Si substantia innominata, strt stria terminalis, Thstv thalamo-striate vein. Note that the neurons of BST have small dimensions, an oval-shaped morphology (often fusiform) and big central clear nuclei with conspicuous nucleoli
On sections passing through the septum, two cell types (slightly pigmented small cells and medium-sized cells with intense staining) are mixed and distributed unevenly.
On sections passing through the anterior commissure, we can observe small medial cells, along the ventricular wall and medium-sized cells on the lateral part of the NBST.
On the most posterior sections, the NBST cells can be easily distinguished from the reticular nucleus cells of the neighbouring thalamus, which are larger and more pigmented. Here again, the medial cells, along the ventricular wall (Nsi contingent), are small and the lateral cells (Nse contingent) are larger. Nse is laterally bordered by prothalamic cell clusters.
On sections concerning the posterior part of the thalamus (level of the lateral geniculate body), the cells are arranged differently, and we can now observe a medial contingent (Nsm) and a paracaudate contingent (Dk), which alone will persist when approaching the amygdala. The Nsm contingent will become increasingly sparse as we approach the end of the caudate nucleus.
Close to the amygdaloid body, the amount of neurons is reduced and may even be minimal. In addition to Nsp cells, whose quantity is also reduced, we can sometimes observe some Nsm cells, of medium size and with an intense coloration. They are arranged ventrally relative to the tail of the caudate nucleus.
H Strenge et al. also included in their work good anatomical diagrams of the sections that he had performed for the BST study. These sections are still used as references for locating the BST. The observations made by H Strenge et al. (1977) were purely histological, even if they could seem, at that time, of little interest. They actually played a major role in the BST study because they showed, for the first time, that the BST had a complex constitution and that we could observe a real zoning with several different cell areas.
Thus, other contributors (JS De Olmos et al. 1985; MM Moga et al. 1989) showed, in mammals, by means of histological and histochemical methods, that there is a cyto-architecture and a chemo-architectony specific to their NBST.
According to these authors, five main sectors were identified in animals, a lateral sector, a medial sector, a central sector, a latero-ventral sector and a postero-medial sector.
The outcome of this research in animals generated a particular enthusiasm, and parallel studies on the human NBST were rapidly conducted. In 1985, P Gaspar et al. carried out immuno-cytochemical studies on the septal region of the human brain and demonstrated its catecholaminergic innervation. The following year, E. Fliers et al. (1986) discovered the presence of vasopressin cells in the BST. Due to the fact that immunohistochemical methods could now be used to locate cell islands with a specific function, this technique was rapidly used to prepare a chemical map of the NBST. The anatomical subdivisions of this formation in humans were thus observed by using these methods, successively by a French team, A Lesur et al. (1989), and then by a German-Hungarian team, A Walter et al. (1991).
According to Lesur et al., there are only four identifiable sectors in humans: the lateral, the central, the medial and the latero-ventral areas. Four markers were used by the authors: synenkephalin, metenkephalin, somatostatin and tyrosine hydroxylase. For the lateral sector, there is no predominance and histochemical markers overlap. The central sector is marked by a “high density of somatostatin neurons”. The medial sector is characterised by “a less dense aminergic and peptidergic innervation”. The latero-ventral sector shows a predominance of the “peptidergic plexi (somatostatin and enkephalin)”.
Walter et al. used a very wide range of histochemical markers (13 in total). In addition to the Lesur markers, they used VIP, substance P, NPH (neurophysins), MBP (myelin basic protein), Cab (calbindin), Syn (synaptophysin), CHR-A (chromogranin), NT (neurotensin), NPY (neuropeptide Y) and the glial fibrillary acid protein. They were able to conclude that there are, in fact, in humans, only three identifiable sectors: the lateral sector, the central sector and the medial sector. The lateral sector, located against the internal capsule, is heterogeneous and includes small neurons and other medium-sized neurons. Most of these neurons are NPY or Syn positive. The central sector consists of a homogeneous set of neurons of similar shape and diameter. This sector has the distinction of being divided into two areas, on coronal sections: a supra-commissural area and a peri-ventral and sub-commissural area. The authors rightly consider that these two areas correspond to the same formation as they all have the same immunohistochemical characteristics: VIP innervation density and presence of somatostatin fibres and neurons (DF Swaab 2003). The medial area is located below the tapered end of the lower edge of the lateral ventricle. It is a heterogeneous sector (such as the lateral sector). It is in this sector that the BST neurons with the smallest diameters are observed. This sector has a VIP innervation (JN Zhou et al. 1995) as well as a less dense peptidergic and aminergic innervation.
Related posts:
 From Prefrontal Lobectomies to Amygdalectomies
Anatomy of Emotions via the History of Neuroanatomy and Neurosciences
From Prefrontal Lobectomies to Amygdalectomies
Anatomy of Emotions via the History of Neuroanatomy and Neurosciences
 Vascularization of the Cerebral Amygdala
Vascularization of the Cerebral Amygdala
 Technique for Dissecting the Amygdaloid Body and Its Close Connections
Technique for Dissecting the Amygdaloid Body and Its Close Connections
 Development of the Human Amygdaloid Complex
Development of the Human Amygdaloid Complex
 The Concept of “Extended Amygdala”
The Concept of “Extended Amygdala”
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


