, Yves Etienne2 and Maurice Niddam3
(1)
Faculté de Médecine, Marseille, France
(2)
Unité de Médecine Légale, Hôpital de la Timone, Marseille, France
(3)
Unité SAMU 13, Centre 15, Hôpital de la Timone, Marseille, France
In 1923, the embryological research and comparative anatomy works performed by JB Johnston revealed the existence, in embryos of lower non-mammalian vertebrates, of grey matter which surrounds a longitudinal association pathway connecting the forebrain and temporal areas. This gave him a glimpse of the existence, in mammals, of a more or less continuous equivalent column of neurons, which follows the stria terminalis, extending from the nucleus accumbens to the amygdala.
JS De Olmos et al. (a team whose major fundamental work concerning the amygdala has already been mentioned) undertook, in 1985, to update this discovery that medical literature had forgotten for over half a century.
Furthermore, the outstanding work of GF Alheid and L Heimer, in 1988, and then of LJ Martin et al. in 1991, detailed and clarified the concept of “the extended amygdala”.
It is to Alheid and Heimer that we owe the English terminology of Extended Amygdala, which shows that this structure functionally originates from the amygdala, of which it is a true extension, and thereby increases its functional capacity.
We can define the extended amygdala as a more or less complete neuronal continuum of small neuronal masses occupying the base of the forebrain from the BST to the central and medial nuclei of the amygdala.
GF Alheid, 2003 and 2009, more accurately, defines the extended amygdala as “a large basal forebrain macrostructure encompassing a contiguous but heterogeneous collection in excess of 16 interconnected and functionally related neuronal aggregates or subnuclei, extending from the centromedial amygdala to the bed nucleus of the stria terminalis”. Such as remarked by GF Alheid, it is important to note that the cortical nucleus, the lateral and the basal complex are excluded from the above definitions of the extended amygdala, despite the fact that it is known that these nuclei project to this structure.
The description of the extended amygdala is complex due to the fact that its content is heterogeneous while it has a relatively homogeneous macroscopic aspect as a long cellular chain connecting the BST and amygdala. It in fact consists of cellular islands and aggregates, which are dispersed rather than clustered, connected by axonal bundles and cellular bridges. Its shape is unique and includes two bends (Figs. 11.1 and 11.2): the first bend occurs between the amygdaloid nuclei and the sub-lenticular basal horizontal portion; the second bend nearly forms a right angle between this horizontal section and the BST, which is positioned sagittally.
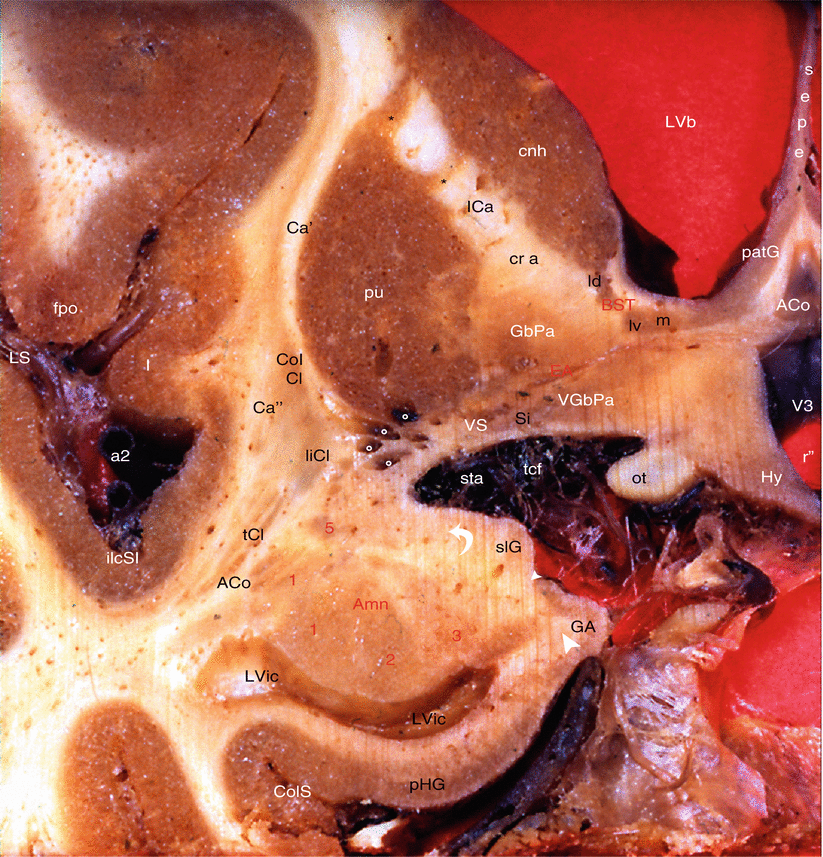
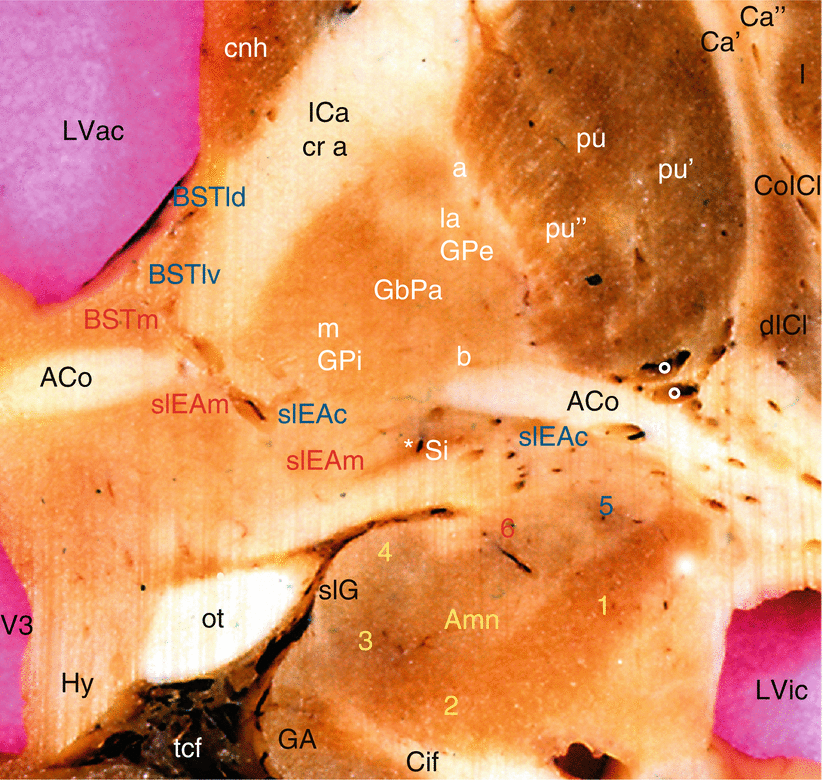
Fig. 11.1
Coronal section of the brain (right half portion) (Through the amygdaloïd nuclear complex and the BST). ACo anterior commissure, Amn (red keys) amygdaloid nuclear complex, 1 lateral, 2 baso-lateral, 3 baso-medial, 5 central, BST bed nucleus of the stria terminalis, ld dorsal division of the pars lateralis, lv ventral division of the pars lateralis, m pars medialis, Ca′ external capsule, Ca″ extreme capsule, cnh caudatus nucleus head, CoICl compact insular claustrum, ColS collateral sulcus, EA extended amygdala, fpo fronto-parietal operculum, GA gyrus ambiens, GbPa globus pallidus, Hy hypothalamus, I insula, ICa (cr a) internal capsule (crus anterior), ilcSI inferior limiting circular sulcus of insula, liCl limitans claustrum, LVb lateral ventricle (body), LVic lateral ventricle (inferior cornu), LS lateral sulcus, ot optic tract, patG paraterminal gyrus, pHG para-hippocampal gyrus, pu putamen, r″ (inf r) infundibular recess, sepe septum pellucidum, Si substantia innominata, slG semi-lunaris gyrus, sta striate artery, tcf transverse cerebral fissure, tCl temporal claustrum, V3 third ventricle, VGbPa ventral globus pallidus, VS ventral striatum, ** putamino-caudate bridges, °° white circles branches of the striate artery, a2 middle cerebral artery, curved white arrow endorhinal sulcus, little white arrowhead semi-annularis sulcus, great white arrowhead sulcus uncal notch
Fig. 11.2
The extended amygdala on a coronal section of the brain through the anterior commissure. Aco anterior commissure, Amn amygdala, 1 lateral nucleus, 2 baso-lateral nucleus, 3 baso-medial nucleus, 4 cortical nucleus, 5 central nucleus, 6 medial nucleus, BSTld dorsal part of the BST’s lateral division, BSTlv ventral part of the BST’s lateral division, BSTm medial division of the BST, Ca′ external capsule, Ca″ extreme capsule, Cif cingulate fasciculus, cnh head of the caudate nucleus, coICl compact insular claustrum, dICl diffuse insular claustrum, GA gyrus ambiens (ambient gyrus), GbPa globus pallidus of the lentiform nucleus; la (GPe: globus pallidus, external part), pars lateralis; m (GPi: globus pallidus, internal part), pars medialis; a external medullary lamina; b internal medullary lamina, a external medullary lamina, b internal medullary lamina, Hy hypothalamus, I insula, ICa internal capsule, cr a crus anterior, LVac lateral ventricle, anterior cornu, LVic lateral ventricle, inferior cornu, ot optic tract, pu putamen, pu′ lateral part of the putamen, pu″ medial part of the putamen, Si substantia innominata, slEAc central division of the sublenticular extended amygdala, slEAm medial division of the sublenticular extended amygdala, slG semi-lunaris gyrus, tcf transverse cerebral fissure, V3 third ventricle, * basal nucleus of Meynert, ° arteries of the putamen; The elements of the central corridor of the extended amygdala are noticed with blue-coloured letters; the elements of the medial corridor of the extended amygdala are noticed with red-coloured letters
The specificity of the extended amygdala is related to the unity of the cellular stream which forms it:
The neurons of the sub-lenticular horizontal portion and those of the central and medial nuclei of the amygdala have the same immunohistochemical characteristics, especially regarding VIP (GM Abrams et al. 1985), neurotensin and above all, angiotensin II (GF Alheid and L Heimer 1988).
Similarly, as recalled by GF Alheid and L Heimer, numerous studies have referred to morphological, histological and histochemical similarities between the BST and the central and medial nuclei of the amygdala (AJ McDonald 1983; G Holstege et al. 1985; JL Price et al. 1987).
We were able, ourselves, to observe the continuity of the extended amygdala components on MRI images of brain sections (see MRI section, etc.).
Similarly, the unity of the extended amygdala appears on sections stained by our Fe Cl3 staining process (see anatomical sections).
Furthermore, we used the tractography process to obtain interesting pictures showing the axonal pathways connecting the BST, the amygdala and the temporal region (Figs. 11.1 and 11.3).
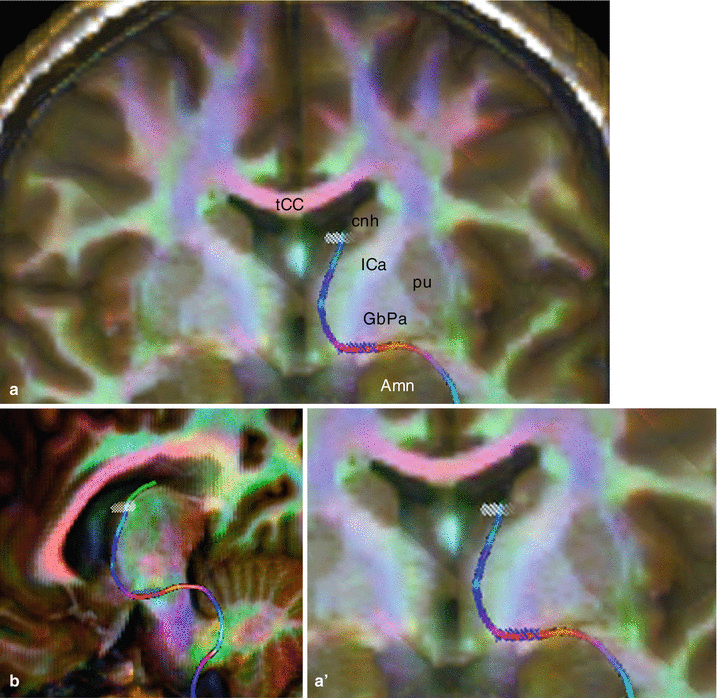
Fig. 11.3
Diffusion Tensor Imaging Fibres Tracking (DTI FT) of the extended amygdala’s pathway. (a) coronal section; (a′) enlargement of a. (b) three quarter view, in order to visualise the change of plan and the two parts of the pathway. Amn amygdaloid nuclear complex, cnh caudate nucleus head, GbPa globus pallidus, ICa internal capsule, pu putamen, tCC truncus of the corpus callosum, white-shaded BST target, blue-shaded Si (substantia innominata) target
It is to be recalled that the basal, horizontal, intermediate, sub-lenticular portion of the extended amygdala is the most complex (JS De Olmos, L Heimer 1999). It is located under the anterior commissure and laterally comprises, near the amygdala, a few elements of the ventral striatum, more medially, the sub-commissural portion of the pallidum or ventral pallidum, and even at the striato-pallidal junction, digitations perpendicular to the axis of the basal portion, which correspond to interpenetrations of the ventral pallidum into the ventral striatum (L Heimer et al. 1991). In addition, at the level of the lower part of the base, elements belonging to the anterior perforated substance mingle with the above-mentioned elements. This complex region, which is crossed by the extended amygdala flow, is what K. Reichert1 has called, for decades, “the innominate substance”2 and that JS De Olmos and L Heimer have developed as a concept: “the concept of the ventral striatopallidal system”.
It is to be recalled that several systems pass through the innominate substance:
Get Clinical Tree app for offline access

The striato-pallidal system interposed between the cell columns that connect the cortex to the brainstem via the thalamus, red nucleus, and substantia nigra (see picture b, Fig. 11.4).
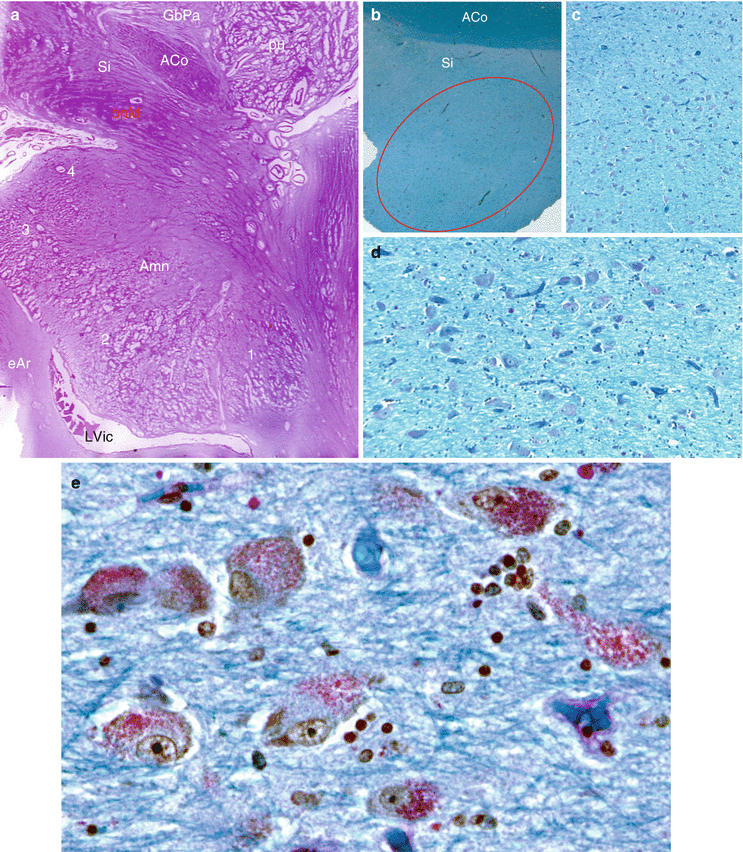
Fig. 11.4
The Basal nucleus or Meynert’s nucleus (a very important component on the route of the extended amygdala). (a) coronal slice of the brain, stained by hematoxylin–eosin, showing the Meynert’s nucleus, situated immediately below the anterior commissure and above the amygdala. (b) enlargement of a, locating the area of the Meynert’s islands in the substantia innominata; (c–e) Luxol fast blue–Pas staining (×100, ×200, ×400) reveals globular-shaped neurons; Their cytoplasm have brown and PAS positive granular material due respectively to their neuromelanin and lipofuschine contents. Aco anterior commissure, Amn amygdaloid nuclear complex, bnM basal nucleus of Meynert, eAr entorhinal area, GbPa globus pallidus, LVic lateral ventricle, inferior cornu, pu putamen, Si Substantia innominata, 1 lateral nucleus of the amygdala, 2 baso-lateral nucleus, 3 baso-medial nucleus, 4 cortical nucleusRelated posts:
 From Prefrontal Lobectomies to Amygdalectomies
Anatomy of Emotions via the History of Neuroanatomy and Neurosciences
From Prefrontal Lobectomies to Amygdalectomies
Anatomy of Emotions via the History of Neuroanatomy and Neurosciences
 Vascularization of the Cerebral Amygdala
Vascularization of the Cerebral Amygdala
 The Bed Nucleus of the Stria Terminalis
The Bed Nucleus of the Stria Terminalis
 Technique for Dissecting the Amygdaloid Body and Its Close Connections
Technique for Dissecting the Amygdaloid Body and Its Close Connections
 Development of the Human Amygdaloid Complex
Development of the Human Amygdaloid Complex

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree

