- •
Myocardial metabolism is inherently and intimately related to cardiac function.
- •
The heart is a metabolic omnivore that uses multiple different substrates for the generation of energy to sustain function.
- •
Nuclear cardiology techniques including SPECT, and especially PET, can be used to measure myocardial metabolism flux in vivo in humans, as well as in animal models. Nuclear imaging techniques are less invasive and have a lower risk than direct arteriovenous catheterization of the heart.
- •
There are several radiotracers that can be used to measure metabolic activity of the heart. 11 C- and 15 O-labeled tracers typically very closely mimic the endogenous substrates that the heart uses and so can be used (in conjunction with compartmental modeling and PET imaging) to quantify not only uptake but also intracellular metabolic processes.
- •
There are several HF risk factors, including obesity, diabetes, hypertension, aging, and ischemia, which each have a typical metabolic signature.
- •
Obesity and diabetes shift the heart toward more fat use, whereas aging, hypertension, and ischemia shift the heart toward more glucose use.
- •
Nonischemic cardiomyopathy and HFrEF typically demonstrate a shift toward relatively more glucose and less fatty acid use.
- •
Although a shift toward glucose use is theoretically advantageous in ischemia, when beta-oxidation of fatty acids is diminished, fat oxidation provides much more ATP/mole than glucose, and so a shift toward glucose metabolism in HFrEF may be maladaptive.
- •
Sex has a major impact on myocardial substrate preference, such that women with nonischemic HFrEF have higher (more normal) fat utilization and blood flow than men with similar ejection fractions. Moreover, women with nonischemic HFrEF typically have a higher survival than men.
- •
Clinicians should be aware that highly metabolically active brown fat can be present in lipomatous hypertrophy of the interatrial septum of the heart. Thus, 18 FDG PET studies that are done for cancer staging can show increased uptake in the heart that is not related to metastasis.
Introduction
Heart failure (HF) is a global pandemic afflicting at least 26 million people worldwide. It is a progressive and mortal disease. Over 75% of patients hospitalized for HF die within 5 years of diagnosis. Unfortunately, the incidence and prevalence of HF are increasing. This increasing burden of HF is driving an increasing interest in myocardial metabolism in HF: how it mechanistically relates to HF; how it may be imaged using nuclear cardiology techniques; how new tracers may be applied; and how myocardial metabolism might be modulated for the benefit of patients with HF. This chapter will delve into some of the answers to these pressing questions and will focus on human studies. It will also seek to illuminate some of these answers with examples of clinical case vignettes and images using nuclear cardiology techniques.
Basic concepts on myocardial metabolism
Myocardial metabolism is inextricably linked to cardiac function. Without substrate metabolism, there would simply be no adenosine triphosphate (ATP) production for systolic and diastolic function. Thus it is critical to understand the fundamentals of human heart metabolism to know how it may be imaged, how it may be deranged in HF, and how it may be leveraged to improve heart function in HF.
The field of myocardial metabolism has evolved rapidly since the early 1950s when Richard Bing and others first discovered that fatty acids are the main fuel of the human heart. The heart, however, is a true “omnivore,” consuming not only fatty acids but also glucose, ketones, lactate, glycogen, triglycerides, and even amino acids to sate its continual need for substrates to metabolize. The heart is able to switch from one fuel to another, a trait known as plasticity , depending on different conditions; substrate availability is key to its ability to keep on beating day after day, decade after decade. The human heart needs to consume incredible amounts of substrates to meet its amazing energy requirements. It is estimated that the human heart consumes around 5 kg/ATP each day or around 2 metric tons per year. This huge energy need helps explain why the adult mammalian heart prefers fatty acid consumption for ATP production. Metabolism of fatty acids, which have more carbons per molecule (e.g., 16 for palmitate, 18 for oleate) compared with glucose (6 carbons) results in greater ATP production per molecule, or per mole. For example, a single palmitate breaks down into acetyl-CoA molecules, which are transformed by the tricarboxylic acid (TCA) cycle and electron transport chain into around 129 ATP. In contrast, full oxidation of glucose results in only around 36 ATP. Nevertheless, oxidation of glucose is approximately 30% more oxygen-efficient than fatty acid oxidation. Thus there is an advantage to glucose use in conditions of limited oxygen, such as ischemia, which is clearly relevant to ischemic cardiomyopathy. There is, however, at least a theoretical disadvantage for the heart to chronically use glucose rather than fatty acids, given the heart’s massive substrate metabolism requirements for generation of ATP.
Methods for measuring substrate metabolism in the human heart
Arteriovenous balance studies
Arteriovenous (A-V) balance studies involve the technique that was pioneered by Bing and colleagues. In this technique, catheters are placed in the coronary arteries and in the coronary sinus. These catheters are then used to directly sample the blood in these locations for measurement of plasma substrates, such as free fatty acids and glucose. The theory is that by measuring how much of each substrate the heart is presented with through the coronary arteries and by subtracting how much of each substrate passes through to the coronary sinus, one can calculate how much of the substrate is taken up and metabolized by the heart. Surprisingly, an early study of myocardial metabolism in patients with HF using this technique did not show much of a perturbation in substrate use. Specifically, a study by Blain et al. in 1956 showed that the failing heart under fasting conditions derived about 58% of its energy requirements from fatty acids, 30% from carbohydrates, 7% from ketone bodies, and 5% from amino acids, results that are similar to the normal human heart. There are several possible explanations for this lack of a difference. Early A-V balance studies did not use concomitant nonradiolabeled tracer infusions (e.g., C-13 palmitate). Using these tracer substrates allows for the distinction between exogenous substrate use and endogenous substrate production. Another possible explanation is that this early study of myocardial metabolism in HF did not include patients with concomitant obesity or diabetes, which would affect plasma substrate levels and hence myocardial metabolism. Indeed, work by Danforth et al. suggested that different etiologies of HF may affect myocardial metabolism differently, an important concept that will be explained later in this chapter. An advantage of A-V balance studies is that direct measurements are made in vivo . A significant disadvantage of this technique is its invasive nature because it requires arterial access and cannulation of the heart’s vessels. Although the absolute risk for a significant complication of A-V balance studies is small, it exists. A-V balance studies are also impractical for longitudinal follow-up in response to pharmacologic or other forms (e.g., bariatric surgery) of interventions.
Magnetic resonance techniques
Magnetic resonance (MR) techniques are predominantly used in the research setting rather than in clinical practice. There are two predominant MR-based techniques: 1 H-MR spectroscopy and the new MR with hyperpolarization and dynamic nuclear polarization (DNP). 1 H-MR specroscopy is used to quantify myocardial fatty acid/triglyceride deposits in the myocardium rather than flux through fatty acid metabolic pathways. In this technique, a region of interest (ROI) is placed on the interventricular septum, so as to avoid contamination of the signal from pericardial/epicardial fat. Because the human heart is made mostly of water, the water peak on the spectra is very large compared with the spectra from fat. Nevertheless, despite the relatively small size of the triglyceride peaks, there is good agreement between 1 H-MR spectroscopy and direct biochemical measurement of lipids in the myocardium. Thus 1 H-MR spectroscopy can be used to evaluate the effect of risk factors for HF, such as obesity and type 2 diabetes, which are associated with steatosis. This technique may also be used to evaluate the effect of different therapies on cardiac steatosis.
MR with hyperpolarization and DNP is a novel technique for measurement of visualization of biochemical pathways such as the Krebs cycle, when 13 C-labeled tracers are used. Hyperpolarization helps improve the signal-to-noise ratio. DNP requires very low temperatures and a high Tesla magnetic field. DNP aligns most nuclear spins, which increases the MR signal. A full review of this new technique is beyond the scope of this chapter but is discussed in detail by Schroeder et al. It should be noted that this technique has only recently been shown to be feasible for evaluating human myocardial metabolism. An advantage of these noninvasive MR-based techniques is the lack of exposure to ionizing radiation. Of course, there are potential downsides with MR techniques, including the high cost, the necessity of a large MR scanner, and the inability to scan patients with certain implanted electronic devices, such as non-MR-compatible implanted cardiac defibrillators (ICDs), which are common in patients with HF with reduced ejection fraction (HFrEF).
Radionuclide techniques
Radionuclide techniques are noninvasive. Unlike MR imaging (MRI), they are not contraindicated in patients with implanted electronic devices such as ICDs and are used for measurement of substrate flux (as opposed to deposition).
Single photon emission computed tomography (SPECT) imaging can be used to evaluate myocardial glucose and fatty acid metabolism. There are several advantages to SPECT imaging, including the general availability of the technology, the simultaneous measurement of cardiac function, and the long half-life of SPECT radiotracers. This long half-life is especially important because it allows SPECT radiotracers to be delivered from an off-site central radiopharmacy rather than requiring that each institution have an on-site cyclotron or generator for radiotracer production. The primary disadvantage of SPECT imaging of myocardial metabolism is that it is only a visual and/or semiquantitative analysis of relative radiotracer uptake in the myocardium. Further limitations of SPECT imaging technology may be seen in its imperfect photon attenuation correction and relatively poor spatial and temporal resolution. That said, SPECT can be used to trace myocardial metabolism.
Table 19.1 shows the primary radiopharmaceuticals used in combination with radionuclide imaging to trace myocardial metabolism. 123 I-β-methyl-P-iodophenylpentadecanoic acid (BMIPP), which is a branched-chain analog of 15-( p -iodophenyl)-pentadecanoic acid (IPPA), is often used in combination with SPECT to estimate fatty acid flux in the myocardium. BMIPP leads to better-quality images than IPPA because the turnover of the latter is so fast. In contrast, the branched chain of BMIPP prevents its β-oxidation and thus increases shunting of the radiotracer into the myocardial triglyceride pools, increasing retention of BMIPP in the heart. This enhances the SPECT images of fatty acid storage (but does not allow for measurement of fatty acid oxidation kinetics). Although glucose metabolism kinetics are not typically measured using SPECT, if used in combination with a specific collimator or detection scheme, the radiolabeled tracer 18 fluorodeoxyglucose (FDG) can be used to measure glucose uptake. Positron emission tomography (PET), however, is used more often than SPECT for measurement of myocardial glucose metabolism.
| Radionuclide | Half-Life | Compound | Present Use |
|---|---|---|---|
| SPECT | |||
| 123 I | 13.3 h | IPPA | Fatty acid uptake, oxidation, and storage |
| BMIPP | Fatty acid storage | ||
| PET | |||
| 15 O | 2.04 min | 15 O 2 | Oxygen consumption |
| 11 C | 20.4 min | Acetate a | Oxygen consumption |
| Palmitate | Fatty acid uptake, oxidation, and storage | ||
| Glucose | Glucose uptake, glycolysis, oxidation, and glycogen turnover | ||
| Lactate | Lactate uptake and oxidation | ||
| Ketone (acetoacetate) | Ketone uptake and oxidation | ||
| OMAR | Imaging of cannabinoid type 1 receptor | ||
| 18 F | 110 min | Deoxyglucose | Glucose uptake |
| FTHA | Fatty acid uptake and oxidation | ||
| FTP | Fatty acid uptake and oxidation | ||
| FCPHA | Fatty acid uptake and oxidation | ||
a 11C-acetate measures tricarboxylic acid cycle and is an indirect index for MVO 2 .
PET has several advantages over SPECT for myocardial metabolism measurement. First, PET can be used to quantify myocardial metabolism processes. Another significant advantage of PET over SPECT is that positron-emitting radionuclides of key elements such as carbon, nitrogen, and oxygen can be used to create tracers that nearly exactly mimic the kinetics of the nonradiolabeled substrates of interest (see Table 19.1 and Fig. 19.1 ). For example, 11 C-palmitate is much closer in structure to palmitate that is not radiolabeled and so 11 C-palmitate can be expected to more closely trace all aspects of fatty acid metabolism than a tracer such as BMIPP, which has a branched side chain that is not a part of the endogenous, unlabeled palmitate. 11 C can be also be used to label acetate, glucose, and lactate to trace the uptake and metabolism of these substrates. 15 O can be used to label acetate to measure total oxygen consumption (MVO 2 ), and 18 F can also be used in place of a hydroxyl group of a substrate/substrate analog of interest without a major alteration of the structure. 18 F is the positron-emitting radionuclide in FDG and in several tracers of fatty acid metabolism (see Table 19.1 ). The main disadvantages of PET are its expense and the complexity of kinetic modeling for quantification of substrate metabolism and of radiotracer generation. Several of the radiotracers (e.g., 11 C-acetate) require an on-site cyclotron for generation, thus limiting their widespread use/distribution.
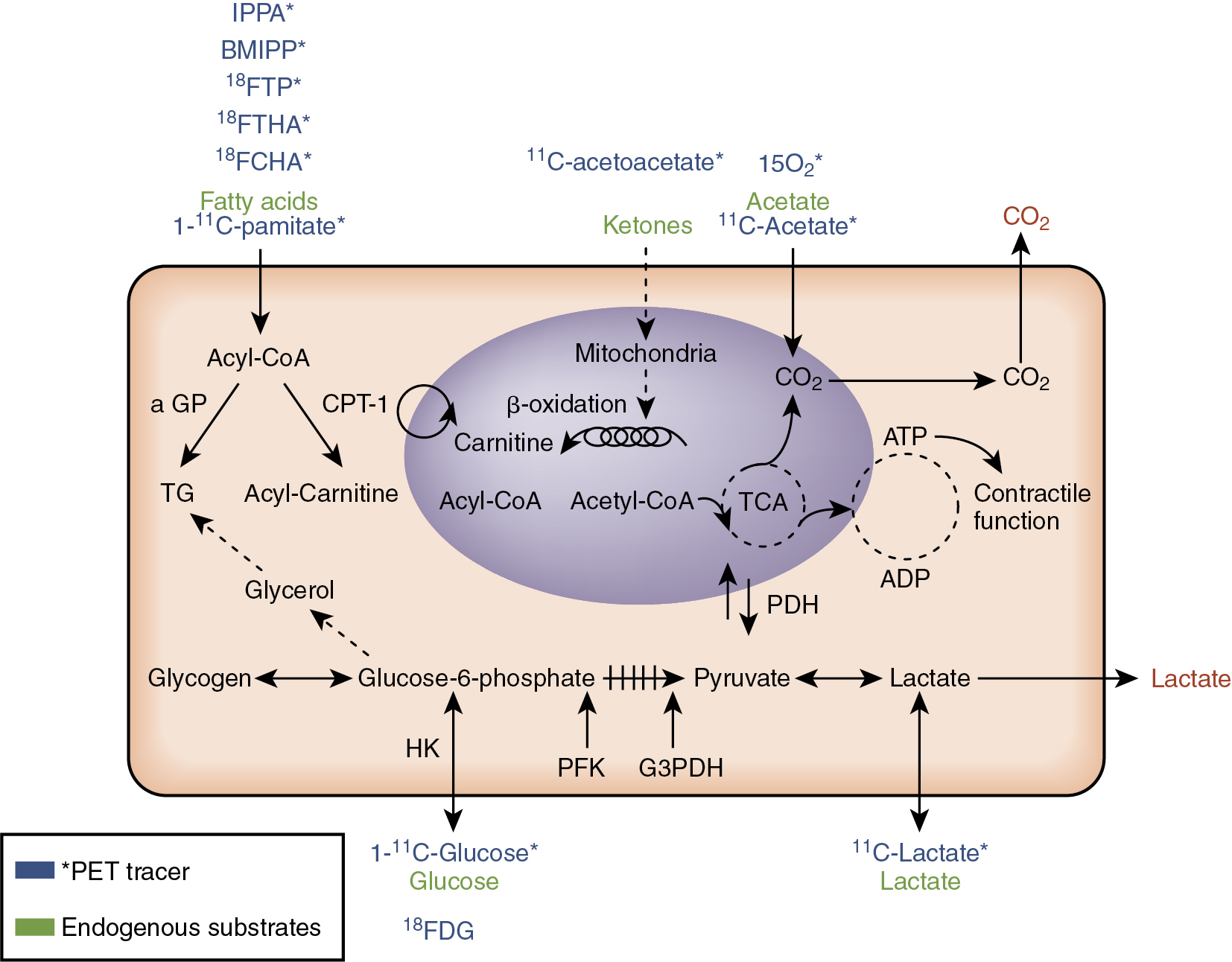
Assessing key myocardial metabolic pathways
PET can be used to quantify uptake and use of several substrates. Although an in-depth review of all possible radiotracers is beyond the scope of this chapter focused on HF metabolism, there are several pathways and radiotracers of particular importance in the assessment of HF and its risk factors.
Myocardial oxygen consumption
As previously mentioned, MVO 2 can be quantified in vivo using 15 O-oxygen or 11 C-acetate. The former has a very short half-life, which lends itself to multiple measurements over a short time period; however, imaging with 15 O-oxygen requires an additional tracer for myocardial blood flow measurement for the quantification of MVO 2 . 11 C-acetate, on the other hand, has around a 20-minute half-life, like all 11 C-acetate compounds. 11 C-acetate is primarily converted to 11 C-acetyl-coA and is metabolized via Krebs cycle. Because the Krebs cycle and oxidative phosphorylation in the mitochondria are closely linked, 11 C-acetate kinetics reflect overall MVO 2 . The quantification of MVO 2 using 11 C-acetate necessitates compartmental modeling or exponential curve fitting. Especially relevant to HF, when there is low cardiac output compartmental modeling, correction for 11 CO 2 in the blood is preferred. This is due to spillover of 11 C activity from the lungs and due to dispersion of the input function.
Carbohydrate metabolism
PET is also used to quantify glucose and lactate metabolism pathways. Measurement of carbohydrate metabolism is especially relevant to HF for several reasons. First, in nonischemic cardiomyopathy and HFrEF, there is a general metabolic shift toward relatively more glucose use. Second, full oxidation of glucose is more oxygen efficient than fatty acid oxidation, which is a theoretical advantage in ischemic cardiomyopathy. Third, identification of viable, glucose-metabolizing myocardium using PET in ischemic cardiomyopathy can alter patient management by helping select those who may benefit from revascularization. The two most commonly used radiotracers of carbohydrate metabolism are 18 FDG and 11 C-glucose. 18 FDG is commonly available, generator-made, and has been used as a radiopharmaceutical for more than 35 years. As discussed in Chapter 20 , 18 FDG is extremely clinically useful for detecting myocardial viability and predicting improved left ventricular (LV) function after revascularization, which is especially relevant to disease management of ischemic cardiomyopathy. 18 FDG is taken up by cardiomyocytes by facilitated transport. The radiotracer is then phosphorylated by hexokinase and trapped in the cell. Thus, 18 FDG cannot be used to study the intramyocellular fate of glucose (e.g., glucose oxidation or glycogen formation), but 18 FDG can be used to assess myocardial glucose uptake and overall flux through glycolysis. To quantify myocardial glucose uptake using 18 FDG, a correction factor, called the lumped constant , must be used to correct for differences between endogenous glucose uptake and the tracer 18 FDG. Unfortunately, the value of the lumped constant may change depending on the substrate and hormonal conditions, which can affect the accuracy of glucose uptake using 18 FDG.
Tracing myocardial glucose metabolism with 11 C-glucose and PET has some advantages over the more commonly used 18 FDG. First, there is no need for a lumped constant because 11 C-glucose closely mimics nonradiolabeled endogenous glucose. Another major advantage is that different intramyocellular fates of glucose (e.g., glucose oxidation, lactate production, glycogen deposition) may be determined using 11 C-glucose tracer kinetics combined with kinetic compartmental modeling. Evaluating the intramyocellular metabolic fate of glucose is especially relevant to myocardial energetics in HF because full glucose oxidation yields around 36 ATP equivalents but glycogen deposition costs around 2 ATP. A significant disadvantage of using 11 C-glucose is its higher cost compared with 18 FDG. The former is also more complicated to synthesize and requires a cyclotron. Moreover, the input function of the kinetic modeling using 11 C-glucose requires correction for 11 CO 2 and 11 C-lactate levels in the input function.
As previously mentioned, 11 C-glucose may be used to measure lactate production, but lactate use by the heart may be measured using the novel PET tracer L-3- 11 C-lactate. The heart is a particularly avid user of lactate when it is available, which is typically during heavy workloads or ischemia. The use of this tracer of lactate metabolism, like 11 C-glucose, requires complex kinetic modeling in conjunction with the PET kinetics. Although lactate metabolism is particularly pertinent to ischemia and hence ischemic cardiomyopathy, this tracer has not been used yet in this condition.
Case vignette 1: Increased cardiac glucose uptake and possible metastasis
A 63-year-old woman with a history of smoking and chronic obstructive pulmonary disease (COPD) presented to an outside hospital for shortness of breath. A chest x-ray revealed a lung nodule that was suspicious for carcinoma. The patient underwent combined computed tomography (CT)/FDG PET, which revealed increased uptake of FDG by the lung mass ( Fig. 19.2 A). Although less distinct, there was also increased FDG uptake by the heart, which suggested a possible cardiac metastasis (see Fig. 19.2 B). The density of the cardiac mass was −57 Hounsfield units (HU) on CT. The patient was transferred to a tertiary care hospital for further evaluation. After transfer, the cardiac mass was further evaluated using transesophageal echocardiography (TEE). This revealed a classic dumbbell-shaped thickening of the interatrial septum on either side of the normal, thin fossa ovalis, consistent with marked lipomatous hypertrophy of the interatrial septum (see Fig. 19.2 C). This was confirmed by cardiac MRI, which revealed the same dumbbell shape, with the signal intensity of the mass being the same as the subcutaneous fat (see Fig. 19.2 D). Additionally, according to steady-state free precession sequences, there was etching at the border of the mass, a fat/water cancellation artifact (not shown).

Lipomatous hypertrophy of the interatrial septum has increased metabolic activity (mimicking cancer) because it is not merely an encapsulated lipoma of the septum but rather can be composed of both brown and white fat cells as well as fibrosis, hypertrophied myocardial cells, with or without chronic inflammatory cells. Brown fat is inherently very metabolically active, especially in cold conditions, because it is involved in thermogenesis. This case illustrates the importance of understanding the possible differential diagnoses for increased cardiac uptake of FDG, other than malignant metastasis. Without an understanding that the FDG uptake was caused by lipomatous hypertrophy of the interatrial septum as opposed to a metastasis, the staging, prognosis, and therapeutic treatment choices for this patient with lung cancer would have been adversely affected. The stage of this patient’s lung cancer would have been changed from a 1 (60% 5-year survival) to a 4 (6% 5-year survival). Thus it is important to recognize that brown fat can exist in adults, specifically in the chest and interatrial septum; FDG uptake is increased by brown fat; lipomatous hypertrophy of the interatrial septum has a distinctive appearance on MRI and TEE; and fat suppression and other techniques can be used to determine that the tissue is indeed fat and not cancer.
Fatty acid metabolism
Both 11 C-palmitate and fatty acid analogs have been used to noninvasively trace myocardial metabolism of the adult heart’s favorite fuel: fat. A primary advantage of 11 C-palmitate for tracing fat metabolism, like 11 C-glucose for tracing glucose metabolism, is that 11 C-palmitate behaves just like the endogenous substrate. Thus, 11 C-palmitate can be used to quantify not only fatty acid uptake but also fatty acid utilization, oxidation, and esterification. Again, increased cost, complexity of production, and the requirement for complex modeling when using 11 C-palmitate are downsides to its use.
Fatty acid analogs, such as 14-(R,S)- 18 F-fluoro-6- thiaheptadecanoic acid (FTHA) and 16- 18 F-fluoro- 4-thia-palmitate (FTP), are alternatives to 11 C-palmitate for tracing fatty acid uptake and metabolism. FTHA is thought to be less reliable for measurement of fatty acid oxidation in hypoxia when fatty acid metabolism should be decreased. FTP, on the other hand, appears to function well as a tracer of uptake in both normoxic and hypoxic conditions. Nevertheless, use of either fluorinated compound requires use of a correction constant, similar to the lumped constant used for glucose metabolism measurement with 18 FDG. Moreover, it is not clear that FTP can be used to accurately quantify fatty acid uptake and oxidation. A relatively new 18 F-labeled tracer, FCPHA trans -9( RS )- 18 F-fluoro-3,4( RS,RS ) methyleneheptadecanoic acid, was created to better trace fatty acid oxidation. Moreover, because beta-oxidation of fatty acid is markedly decreased in ischemia, 18 FCPHA has recently been shown to be a sensitive marker of ischemia in patients with coronary artery disease (CAD). In a report of the first comparison of rest-stress 18 FCPHA (CardioPET) versus standard corrected stress-rest 99m technetium (Tc)-sestamibi myocardial perfusion using SPECT, the former appeared to more closely correlate with the extent of ischemia predicted by the catheterization than the sestamibi images on SPECT. This has obvious implications regarding the evaluation of patients with ischemic cardiomyopathy.
Ketones, which in mammals are predominantly derived from fat oxidation in the liver, are another important substrate for myocardial use. Ketone generation is increased in physiologic states, such as fasting, and in pathophysiologic states, such as diabetic ketoacidosis. Ketones are not only fuels for organs such as the heart, skeletal muscle, and brain, but they are also important in signaling, modification of proteins, oxidative stress, and inflammation. This chapter will focus on ketones as a substrate for myocardial metabolism. Although the heart “cannot live on ketones alone” because it cannot replenish citric acid cycle intermediates through anaplerosis, ketones remain an important substrate, especially in fasting states and after prolonged exercise. Ketones can be converted into acetyl CoA, which can then be oxidized in the Krebs cycle. There are also recent studies that show that ketogenic diets and exogenous ketones in a ketone monoester have benefits on physical performance in healthy adults. Whether ketones are particularly beneficial for patients with HF is not completely understood. Nevertheless, plasma ketones can be increased with HF, and there is a new PET tracer of ketone uptake, 11 C-acetoacetate, which should help in the evaluation of myocardial ketone use in vivo in humans.
Case vignette 2: Decreased myocardial fat uptake with lipoprotein lipase deficiency
A 32-year-old white woman of French-Canadian ancestry with a history of lipoprotein lipase (LPL) deficiency was seen in routine follow-up. She had a past surgical history of a cholecystectomy. She also had a history of pancreatitis while on oral contraceptive therapy, which was when her LPL deficiency was first diagnosed. Her peak plasma triglyceride level was 7000 mg/dL. She has been treated with both fish oil and with fenofibrate therapy. She had one more bout of pancreatitis and subsequently changed her diet, incorporating medium-chain triglyceride oil and increasing her exercise. She underwent PET imaging with 15 O-water, 18 FDG, and 11 C-palmitate both at rest and with low-dose dobutamine (5 mcg/kg/min) to simulate exercise. The patient’s myocardial blood flow was normal at rest and increased mildly with dobutamine (0.94 and 1.05 mL/g/min, respectively). Glucose utilization in the anterolateral wall was relatively low at 19.6 nmol/g/min at rest in the fasting state ( Fig. 19.3 , left top panel); however, it markedly increased with even low-dose dobutamine to 75.8 nmol/g/min (see Fig. 19.3 right top panel). There was an increase in plasma glucose at the time of the scans (from 68 at rest to 78 mg/dL on dobutamine), which likely aided glucose utilization slightly. In contrast to the picture of glucose metabolism, myocardial fatty acid utilization and oxidation barely changed after dobutamine stimulation (see Fig. 19.3 , bottom panels). Congruent with this, plasma fatty acid levels were nearly identical at rest and on dobutamine (0.23 and 0.24 meq/L). This lack of increase in myocardial fatty acid metabolism in this case is in stark contrast to the normal increase seen in healthy young controls without LPL deficiency. This case illustrates the important points that plasma substrate availability influences myocardial substrate metabolism and that inborn errors of fatty acid metabolism (e.g., LPL deficiency) detrimentally affect the adult heart from upregulating use of its generally preferred substrate (fatty acids) after stimulation with dobutamine (or exercise).

Related posts:
 Radiopharmaceuticals for clinical SPECT and PET and imaging protocols
Radiopharmaceuticals for clinical SPECT and PET and imaging protocols
 Imaging in patients with acute chest pain in the emergency department
Imaging in patients with acute chest pain in the emergency department
 Key concepts in risk stratification and cost-effectiveness using nuclear scintigraphy in stable coronary artery disease
Key concepts in risk stratification and cost-effectiveness using nuclear scintigraphy in stable coronary artery disease
 Molecular imaging of myocardial infarction and remodeling
Molecular imaging of myocardial infarction and remodeling
 Artificial intelligence in nuclear cardiology
Artificial intelligence in nuclear cardiology
 Large-vessel vasculitis
Large-vessel vasculitis
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


