Chapter 12 Embryology and development
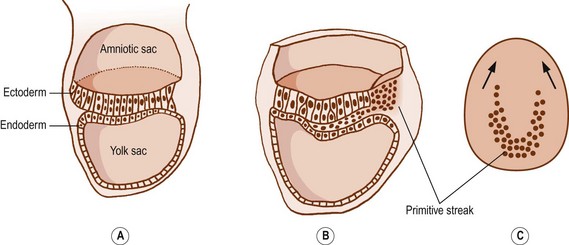
After 15 days of development, the human embryo is in the form of a flat, ovoid disc which consists of two layers of cells: the ectoderm dorsally and the endoderm ventrally (Fig. 12.1). The ectoderm is that layer which principally will give rise to the skin and spinal cord. The endoderm forms the alimentary tract.1 At the caudal end of the embryo, the cells of the ectoderm become rounded and heap up, forming an elevation known as the primitive streak.1 Cells from the primitive streak migrate laterally and forwards, insinuating between the ectoderm and endoderm to form a third layer in the embryo called the mesoderm (Figs. 12.1, 12.2). Just in front of the primitive streak, another thickening develops, known as Hensen’s node. From this node, a cord of cells, known as the notochord, migrates forwards between the ectoderm and endoderm (see Fig. 12.2). By about 28 days, the notochord fully demarcates the midline of the embryo1 and induces the formation of the vertebral column around it. Dorsal to the notochord, the ectoderm forms the neural tube, which differentiates into the brain and spinal cord.
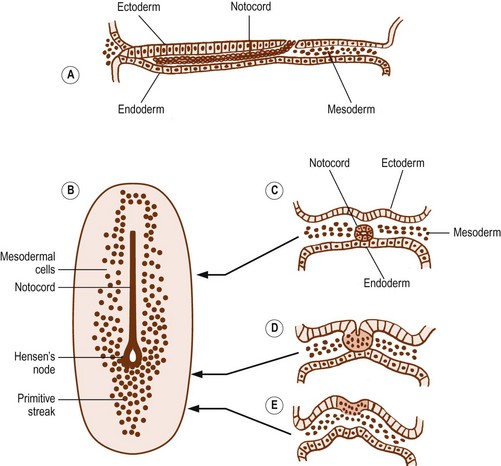
On each side of the notochord, the mesoderm of the embryo is thickened to form a longitudinal mass known as the paraxial mesoderm. By the 21st day of development, the paraxial mesoderm starts to be marked by transverse clefts across its dorsal surface. These clefts separate the paraxial mesoderm into segments called somites (Fig. 12.3). The first somites appear in the region of the head, and others appear successively caudally. By about the 30th day of embryonic development, a total of 42–44 somites are formed.1
The clefts demarcating the somites are actually indentations, so the segmentation they create is apparent only along the dorsal aspect of the paraxial mesoderm. Deeply, beneath the surface of the embryo, the paraxial mesoderm remains a single, longitudinally continuous mass.2 Using the transverse clefts as a guide, however, the further development of each somite can be traced.
The 42–44 somites of the human embryo can be named as 4 occipital, 8 cervical, 12 thoracic, 5 lumbar, 5 sacral and 8–10 coccygeal. The first occipital and the last 7–8 coccygeal somites regress and give rise to no permanent structures.1 The remaining three occipital somites are involved in the formation of the occipital region of the skull and the tongue. The other somites form the vertebral column and the trunk.
The cells in the somites are originally epithelial in nature but they gradually change into loosely arranged tissue called mesenchyme (Fig. 12.4). In transverse section, each somite is roughly triangular in outline, presenting ventral and dorsolateral borders, and a medial border facing the neural tube (Fig. 12.5).
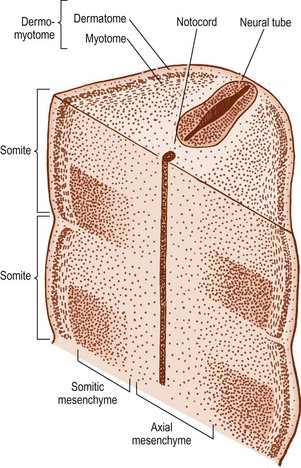
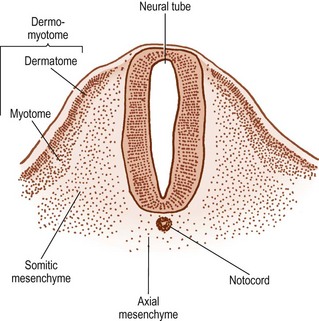
Within the somite, two clusters of cells develop. Those cells in the ventral and medial regions of the somite rapidly multiply and form a mass, which, in the past, has been referred to as the sclerotome, but for reasons outlined elsewhere2 the term somitic mesenchyme is used here. These cells are exclusively involved in the formation of the vertebral column. The remaining cells, along the dorsolateral border of the somite, give rise to the musculature and skin of the trunk and are collectively referred to as the dermomyotome.
The fate of the somitic mesenchyme
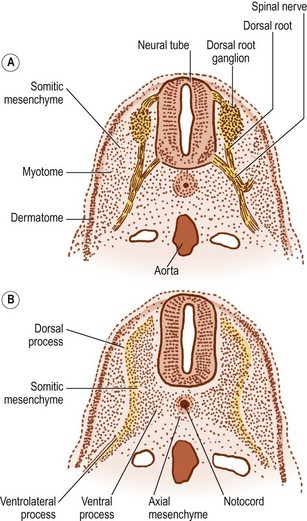
The notochord lies between the aorta ventrally, and the neural tube dorsally. The neural tube is flanked by the somitic mesenchyme, but the somitic mesenchyme initially does not extend as far medially as the notochord. The notochord is surrounded separately by a continuous column of very loose-meshed mesenchyme called the axial mesenchyme (see Fig. 12.5).2 The density of the axial mesenchyme gradually increases as these cells multiply and surround the notochord (Figs. 12.6, 12.7). Meanwhile, a separate series of events occurs in the somitic mesenchyme.
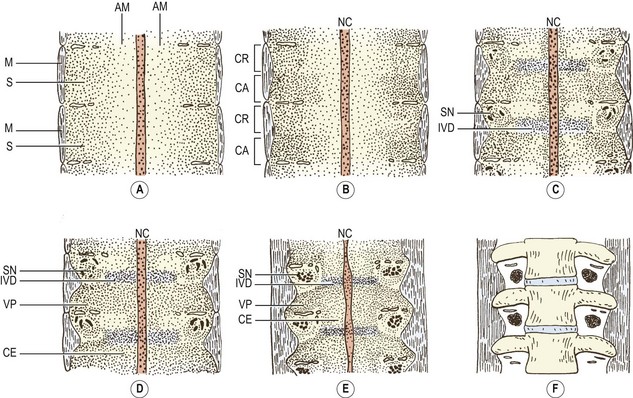
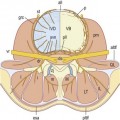
In the caudal half of each somite, the density of nuclei increases, giving it a darker staining appearance (see Figs 12.4 and 12.7B). The cranial half of the somite remains less dense and is invaded by the developing spinal nerve (see Figs 12.6A and 12.7C). The nerve grows laterally to invade the dermomyotome, and as the nerve increases in length and thickness, the cells of the cranial half of the somite come to be arranged in concentric layers around the nerve.2 In time, the developing nerve occupies most of the entire cranial half of the somite, which itself gives rise to little but perineural tissue. It is the denser, caudal half of each somite that participates in the formation of the vertebral column.
In the caudal half of each somite, two processes develop: a dorsal process and a ventrolateral process.2 The dorsal process spreads dorsally to surround the neural tube and will give rise to the neural arch (see Fig. 12.6B). Hence, it is also referred to as the arcual process. The ventrolateral process extends laterally and gives rise to the costal element of the future vertebrae. Hence, it is also referred to as the costal process (see Fig. 12.6B). In the lumbar region, the costal elements of each vertebra are represented in the form of the transverse processes.
As the axial mesenchyme increases in density, its cells assume a concentric orientation around the notochord. These cells will form the greater part of the future vertebral body, and the portion of the body that they form is referred to as the centrum (see Fig. 12.7). Opposite the lower half of the cranial portion of the adjacent somite, a zone of higher density develops in the axial mesenchyme (see Fig. 12.7). This zone forms the predecessor to the future intervertebral disc.2
While these events take place in the axial mesenchyme, a third process develops in the somitic mesenchyme. This process, known as the ventral or chordal process, extends towards the notochord to blend with the axial mesenchyme just caudal to the zone of the future intervertebral disc.2 In this way, the chordal process connects the somitic mesenchyme with the centrum of the vertebral body, and the vertebral body is eventually formed by the centrum and the terminal portions of the chordal processes from each side.
The dorsal processes of the somitic mesenchyme continue to extend around the sides of the neural tube, and just lateral to the developing dorsal root ganglion, the dorsal processes of adjacent somites blend with one another at the sites of the future zygapophysial joints.2 Elsewhere, the neural arches of adjacent segments are bridged by less dense condensations of mesenchyme that will give rise to the ligaments of the neural arch.
By this stage of development, the shape of the future vertebra is outlined by mesenchymal tissue. Condensations of the axial mesenchyme have surrounded the notochord and have moulded the vertebral body. The future intervertebral disc has condensed in the axial mesenchyme opposite the lower half of the cranial portion of the somitic mesenchyme. The cranial half of each somite has condensed around the developing spinal nerve and will form only perineural tissue. The condensed caudal half of the somitic mesenchyme has formed three processes. A ventral process blends with the axial mesenchyme below the intervertebral disc, while a dorsal process embraces the side of the neural tube. Together, the ventral and dorsal processes outline the future neural arch. The ventrolateral process radiates from the neural arch on each side to outline the future transverse process. At this stage of development, the left and right dorsal processes do not yet meet behind the neural tube and are united only by a membrane.3,4 The neural arch is completed dorsally at a later stage of development.
The fate of the dermomyotome
Initially, two types of cells are evident in the dermomyotome. Epithelial cells cover the dorsolateral surface of the somite and can be recognised as the dermatome. Deep to these lie mesenchymal cells, collectively known as the myotome. Gradually, the cells of the dermatome lose their epithelial character and become incorporated into the myotomal mass, but they remain attached to the overlying ectoderm and give rise to the dermis and subcutaneous tissues.1 The cells of the myotome give rise to muscular tissue.
The myotomal mass maintains its ventrolateral location in relation to the somitic mesenchyme. Opposite the condensed caudal half of the somite it is gradually displaced laterally by the developing ventrolateral process. Opposite the looser cranial half of the somite, it bulges towards the somite but is also indented by the developing spinal nerve (see Fig. 12.7).2
As the spinal nerve divides into a ventral and dorsal ramus at about the 40th day of development,4 the myotome splits into two portions.1 The division occurs along a plane depicted by the developing transverse processes, and the two portions are separated by a septum that forms the future intertransverse ligaments (Fig. 12.8). The dorsal portion of the myotome is known as the epimere, or epaxial portion, and is innervated by the dorsal ramus of the spinal nerve. The ventral portion is known as the hypomere, or hypaxial portion, and is innervated by the ventral ramus of the spinal nerve.
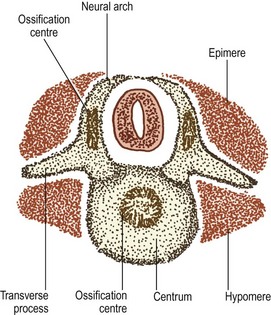
The epimeres throughout the vertebral column divide further into medial and lateral divisions,1,5 which are supplied by the medial and lateral branches of the dorsal rami, respectively. In the lumbar region, the medial division forms the multifidus muscle, while the lateral division forms the iliocostalis and longissimus muscles.
Chondrification
As the mesenchymal models of the vertebrae are being completed, some of the mesenchymal cells change character and become cartilaginous. This occurs at about the 6th week of gestation4 and heralds the onset of the cartilaginous phase of vertebral development.
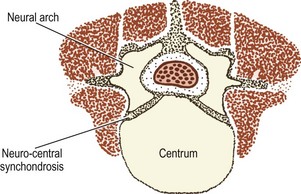
A pair of chondrification centres appear in the centrum of each vertebra. They rapidly fuse into one centre, which expands to chondrify the entire centrum.1 Chondrification centres also appear in each half of the neural arch. These expand dorsally through the dorsal process of the somitic mesenchyme on each side, and meet one another behind the neural tube to complete the neural arch. From the site of union, a cartilaginous spinous process develops dorsally. The neural arch centres also extend laterally to chondrify the transverse process, and ventrally along the ventral process of the somitic mesenchyme to blend with the chondrifying centrum.
As a consequence of these events, a cartilaginous model of the future vertebra is laid down, but even as chondrification of the vertebral column is being completed, these cartilaginous models start to be replaced by definitive, osseous vertebrae (see Fig. 12.8).
Ossification
Ossification is the third phase of development of the vertebral column. It commences during the 9th–10th weeks of intrauterine life,6 but is not completed until adolescent life. The first process of ossification is called primary ossification and occurs at sites where blood vessels invade the cartilaginous models of the future vertebrae.
The cartilaginous neural arches are invaded from behind to form a primary ossification centre in each half of the neural arch (see Fig. 12.8). The cartilaginous vertebral body is invaded by blood vessels through its anterior and posterior surfaces. Some authorities maintain that these two sets of blood vessels give rise, respectively, to separate ventral and dorsal ossification centres, which rapidly fuse to form a single ossification centre in the middle of the future vertebral body,7 but others maintain that this phenomenon is only a variation that occurs in about 5% of cases.8,9 Another variant is to have two centres lying lateral to one another8 but the most common pattern is to have one single centre.8
The onset of ossification differs according to vertebral level and the part to be ossified. Primary ossification centres in the neural arches first appear at cervicothoracic levels, followed by upper cervical and then thoracolumbar levels. Centres in the neural arches then appear progressively in cranial and caudal directions from these levels.10 Primary centres in the vertebral bodies first appear at lower thoracic and upper lumbar levels, and then progressively appear at levels above and below these.10 In this way, ossification centres are established in the bodies and neural arches of the lumbar vertebrae by the 12th–14th week of gestation.
In the centrum of the vertebral body, the primary ossification centre expands radially and towards the intervertebral discs above and below. It reaches the anterior aspect of the centrum by about 22 weeks of antenatal life, and the posterior aspect by about 25 weeks,11 but ossification does not reach the superior and inferior surfaces of the vertebral body, which remain cartilaginous and form the growth plates of the vertebral body. In the neural arches, ossification extends in all directions from the primary centre: ventrally towards the vertebral body; laterally into the transverse process; and dorsally around the neural tube.
At birth, the lumbar vertebrae are still not completely ossified (Figs. 12.9, 12.10). The bulk of the centrum is ossified, and in lateral radiographs has the appearance of an ovoid block of bone with convex upper and lower surfaces.12–14 Large vascular channels penetrate the anterior and posterior aspects of the centrum,7 and on radiographs of neonatal spines these appear as areas of translucency.14 The upper and lower surfaces of the vertebral body are still covered by the thick cartilage plates, and the combined height of these plates and the intervertebral disc is approximately the same as the height of the ossified lumbar vertebral bodies.12–14 The pedicles and the proximal parts of the laminae and transverse processes are ossified but the spinous processes and the distal parts of the transverse processes are still cartilaginous. The articular processes are ossified for the most part but their distal ends remain cartilaginous.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


