Anatomy of the Brain
The advent of high-resolution computed tomography (CT) and magnetic resonance imaging (MRI) scanners has allowed the fine anatomic structure to be seen in detail. In this chapter the gross surface anatomy of the brain is briefly reviewed, followed by discussion of CT and MRI techniques, followed by the illustration of sectional anatomy in multiplanar axes in these two modalities.
Overview of Brain Anatomy
The brain is semisolid and conforms to the shape of the skull. Its hemispheric surface is convoluted and has gyri and sulci. The brain consists of the cerebrum, cerebellum, and brainstem.
Dura and Dural Structures
The brain is protected (from the outer to the inner layer) by the skull, meninges, and cerebrospinal fluid (CSF). The meninges are organized into dura mater, arachnoid membrane, and pia mater, in order of their proximity to the skull. The pia mater follows all the gyri and is separated from the arachnoid membrane by CSF. The potential subdural space in turn separates the arachnoid membrane from the dura. The outer parietal layer of the dura adheres to the skull. The dural venous sinuses are made between the dural reflections and their opposing edges, thereby forming the superior and interior sagittal, transverse, sigmoid, cavernous, and straight sinuses. These sinuses drain to the jugular venous system.
Ventricles
There are four ventricles within the brain. They are lined with ependyma and contain the CSF, produced by the choroid plexus.
Lateral (Left and Right) Ventricles.
Lateral ventricles are formed by the two ependyma-lined cavities of the cerebral hemisphere and communicate with the third ventricle via the midline foramen of Monro ( Fig. 8-1 ). They can be divided into five parts: the anterior (frontal) horn, the ventricular body, the collateral (atrium) trigone, the inferior (temporal) horn, and the posterior (occipital) horn.
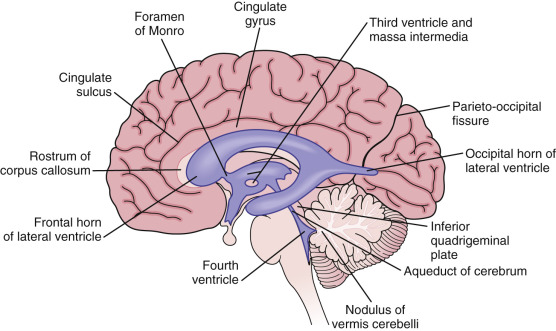
Third Ventricle.
The third ventricle is a slitlike ventricle midline cavity of the diencephalons. It communicates with the lateral ventricles via the interventricular foramina of Monro and with the fourth ventricle via the cerebral aqueduct. It contains choroid plexus in its roof.
Fourth Ventricle.
The fourth ventricle is a rhomboid-shaped cavity overlying the pons and medulla, extending from the central canal of the upper cervical spinal cord to the cerebral aqueduct of the midbrain. There is a small lateral recess on each side of the fourth ventricle that contains choroid plexus that protrudes through the foramina of Luschka into the subarachnoid space. A small median aperture in the caudal part of the ventricle is known as the foramen of Magendie. Via the two lateral foramina of Luschka and the single medial foramen of Magendie, CSF flows into the ventricular system into the subarachnoid spaces.
Cerebral Hemispheres
The two cerebral hemispheres are separated by interhemispheric fissures and falx cerebri. On the lateral surface of the brain, the sylvian fissure (lateral fissure) and the rolandic fissure (central fissure) separate the cerebral hemisphere into the frontal lobe, temporal lobe, parietal lobe, and a line drawn from the parietooccipital sulcus onto the preoccipital notch, delineating the boundaries of the parietal and temporal lobes from that of the occipital lobe.
The frontal lobe, the largest of all the brain, has four principal gyri: the precentral gyrus and the superior frontal, middle frontal, and inferior frontal gyri . The precentral gyrus, parallel to the central sulcus, together with the anterior bank of the central sulcus comprises the primary motor area, which is one of the most important cortical areas for movement. Rostral to the precentral sulcus is the premotor area, another important area for movement. The middle frontal gyrus contains Brodmann’s area 8, known as the frontal eye field, which is important for conjugate eye movements. Another important motor area for speech, Broca’s area, is located at the triangular and opercular parts of the inferior frontal gyrus in the dominant hemisphere.
In the parietal lobe there are a postcentral gyrus, a superior parietal lobule, and an inferior parietal lobule. The postcentral gyrus is a primary somesthetic area involved in general body sensation. The superior parietal lobule is involved with the behavioral interaction of an individual with the surrounding space, and the inferior parietal lobule with the integration of diverse sensory information for speech and perception.
In the temporal lobe the superior, middle, and inferior temporal gyri are separated by the two transverse sulci. The superior temporal gyrus contains two important functional structures. One is the transverse temporal gyri of Heschl, which constitutes the primary auditory area, and another is Wernicke’s area caudal to the transverse gyri of Heschl, which is involved in the comprehension of spoken language. The inferior temporal gyrus is involved with perception of visual form and color. In the occipital lobe the occipital pole contains a portion of the primary visual area, which is more extensive on the medial aspect of the occipital lobe.
On the medial surface of the hemisphere, most convolutions are somewhat flat, but the corpus callosum is the most prominent structure; it connects the two hemispheres and plays an important role in the transfer of information between the two hemispheres. Ventral to the corpus callosum is the septum pellucidum, which separates the two lateral ventricles. Medially the paracentral lobule represents medial extensions of the precentral and postcentral gyri. Caudal to the paracentral lobule is the precuneus, which lies immediately rostral to the parietooccipital sulcus. The parietooccipital sulcus separates the parietal and temporal lobes from the occipital lobe. The occipital lobe is divided into the lingual and the cuneus gyri by the calcarine sulcus. On each bank of the calcarine sulcus is the primary visual area. Between the medial aspect of the temporal lobe and the lateral aspect of the midbrain is the hippocampal gyrus, which mediates verbal learning and memory. The hippocampal sulcus separates the hippocampal gyrus from the parahippocampal gyrus. The parahippocampal gyrus in turn is separated from the fusiform gyrus by the collateral sulcus and terminates in the uncus. The medial surface of the cerebral hemisphere is superior to the delineating components of the limbic lobe, such as the subcallosal gyrus, cingulated gyrus, parahippocampal gyrus, and uncus.
Hemispheric White Matter
The massive white matter contains basically three types of fibers: projection fibers, association fibers, and commissural fibers. The projection fibers convey impulses from the cortex to the distant loci or vice versa. The association fibers connect the adjacent cortex within the same hemisphere. The commissural fibers connect corresponding cortex of the two hemispheres.
The projection fibers are the following:
- •
Corona radiata—convey impulses to and from the cortex, enter the projection fibers arranged as radiating bundles. Near the upper part of the brainstem these fibers form the internal capsule.
- •
Optic radiation (also known as the geniculocalcarine tract or the geniculostriate pathway )—the fibers arise from the most posterior component of the posterior limb of the internal capsule and radiate toward the calcarine sulcus.
- •
Thalamocortical radiation—arises mainly from the thalamus and projects to nearly all regions of the cortex
The intrahemispheric commissures (association fibers) are the following:
- •
Uncinate fasciculus—connect the orbital frontal gyri and parts of the inferior and middle frontal gyri to anterior portions of the temporal lobe
- •
Arcuate fasciculus—connect the superior and middle frontal gyri to parts of the temporal lobe
- •
Cingulate fibers—connect the frontal and parietal lobes to the parahippocampal gyrus and adjacent temporal gyrus
- •
Superior longitudinal fasciculus—connect the occipital lobes to the parietal and frontal lobes
- •
Visual radiations—connect the lateral geniculate bodies to the occipital lobes
- •
Fornix—connects the hippocampus to the ipsilateral mammillary body
The interhemispheric commissures (commissural fibers) are the following:
- •
Anterior commissure—connect the temporal lobes and olfactory structures on the two sides
- •
Posterior commissure—connect the rostral midbrain nuclei, located behind the third ventricle
- •
Corpus callosum—connect all corresponding cortex of the opposite hemisphere
Posterior Fossa
The posterior fossa contains the cerebellum and brainstem. The posterior fossa is outlined by the clivus and petrous bones anteroinferiorly, the tentorium cerebelli superiorly, and the occipital bone posteroinferiorly. The cerebellum is located posteriorly in the two thirds of the posterior fossa, separated from the brainstem by the fourth ventricle. The brainstem occupies the anterior third of the posterior fossa, including the midbrain, pons, and medulla oblongata.
The midbrain consists of cerebral peduncles anteriorly and tectum posteriorly. The prominent cerebral peduncles leave a triangular depressed area called the interpeduncular fossa . The basilar artery lies in this fossa and the third cranial nerve emerges from it. The dorsal surface of the midbrain is composed of the tectum and tegmentum. The tectum represents the superior and inferior colliculi. Caudal to the inferior colliculus, the fourth cranial nerve, the only cranial nerve, exits from the dorsal aspect of the brainstem. Dorsal to the tegmentum is the cerebral aqueduct, connecting the third and fourth ventricles.
The pons on the ventral surface is separated from the crus cerebri by the superior pontine sulcus and from the medulla by the inferior pontine sulcus. The massive transverse fibers in the ventral portion of the pons form the middle cerebellar peduncle. On the dorsal surface the pontine tegmentum is continuous with the reticular formation of the medulla and midbrain. Laterally the cerebellopontine angle (CPA) is formed by the junction of pons, medulla, and cerebellum. From the CPA emerge the seventh and eighth cranial nerves. Caudally the ninth and tenth cranial nerves cross the CPA; superiorly the fifth nerve crosses it.
The medulla is the most caudal portion of the brainstem and continuous with the cervical cord at the level of the foramen magnum. Anteriorly the pyramid has a protrusive appearance formed largely by the corticospinal tracts. Posterior and lateral to the pyramid is a very shallow groove called the ventrolateral sulcus in which the vertebral arteries usually lie. Lateral to the pyramid is the olive, corresponding to the inferior olivary nucleus. Ventrolaterally the ninth, tenth, and eleventh cranial nerves emerge from the postolivary sulcus, and the twelfth cranial nerve emerges from the preolivary sulcus. Laterally the inferior cerebellar peduncles, connecting the spinal cord and medulla with the cerebellum, make the superior protrusion.
The cerebellum is attached to the brainstem by three cerebellar peduncles and forms the roof of the fourth ventricle. It consists of the vermis and hemispheres. The two cerebellar hemispheres are united by the vermis as a midline structure. On the inferior surface, two deep sulci clearly separate the vermis from the hemispheres. The cerebellum is divided by the primary fissure into anterior and posterior lobes and is divided by the horizontal fissure into superior and inferior halves.
Vascular Supply
The brain derives its vascular supply ( Fig. 8-2 ) from two carotid and two vertebral arteries. The internal carotid artery bifurcates terminally into the anterior and middle cerebral arteries. The two vertebral arteries unite at the caudal border of the pons to form the basilar artery. Anterior to the interpeduncular fossa, the basilar artery divides into the two posterior cerebral arteries. Anterior and posterior communicating arteries and proximal portions of the anterior, middle, and posterior cerebral arteries form the cerebral arterial circle called the circle of Willis .

The anterior cerebral artery supplies blood mainly to the frontal lobe medially and mediolaterally, with some supply to the parasagittal parietal lobe. The medial lenticulostriate arteries, arising from the horizontal portion of the anterior cerebral artery, supply the anterior aspects of the putamen and caudate nucleus, the anteroinferior part of the internal capsule, and the dorsal aspect of the chiasm and the hypothalamus. The pericallosal branches supply the upper surface of the corpus callosum and septum pellucidum. The medial surface of the hemisphere is supplied by the hemispheric branches of the anterior cerebral artery. The anterior choroidal branch of the internal carotid artery supplies the remaining portion of the internal capsule as well as the choroid plexus. The middle cerebral artery, the largest of the cerebral arteries, gives off five to eight branches into the insular region called the sylvian triangle and branches from the lateral sulcus over the lateral convexity of the hemisphere. These branches supply blood mainly to the frontal lobe inferiorly and laterally, most of the temporal lobe, the temporooccipital and angular areas, and the motor and premotor areas of the parietal lobe. The lenticulostriate arteries, the first branches to arise from the horizontal portion of the middle cerebral artery, supply the lateral basal ganglia. The posterior cerebral artery divides mainly into the posterior temporal artery and the internal occipital arteries. The former supplies the posterior temporal lobe and the occipitotemporal and lingual gyri; the latter supplies the occipital lobe, portions of the splenium of the corpus callosum, and part of the temporal lobe.
The anterior cerebral territory is located between the anterior part of the interhemispheric fissure and a line along the anterior lateral wall of the lateral ventricle, and the posterior cerebral territory is located between the posterior portion of the interhemispheric fissure and a line along the posterior lateral wall of the lateral ventricle. The area between the lines along the lateral ventricle wall is the middle cerebral territory. The ill-defined anterior area between the anterior cerebral artery and the middle cerebral artery is called the anterior watershed region. The area between the middle cerebral and posterior cerebral arteries is the posterior watershed area. These two areas are more susceptible to hypotensive infarctions than other areas of the brain are.
In the posterior fossa, each half of the cerebellum is supplied by the posterior inferior cerebellar artery, the anterior inferior cerebellar artery, and the superior cerebellar artery. The posterior inferior cerebellar artery, derived from the vertebral artery, supplies the inferior vermis, inferior cerebellar hemispheres, and choroid plexus of the fourth ventricle. The anterior inferior cerebellar arteries, arising from the caudal part of the basilar artery, supply mainly the area of the CPA and the anterior inferior cerebellar hemispheres. The perforator branches of the basilar artery supply the pons. The superior cerebellar arteries, arising from the rostral part of the basilar artery, supply the superior portion of the cerebellar hemispheres.
CT Technique
Axial head CT scanning is performed traditionally with the angulation of the gantry parallel to the orbitomeatal line, defined as passing through the lateral canthus and middle of the external ear canal. To avoid radiation through the eyes when performing axial head CT, a 15- to 20-degree angulation of the gantry to the canthomeatal line is advisable. To match the MRI slices, axial head CT scanning is advocated to parallel the Talairach anterior commissure–posterior commissure (AC-PC) line with the angle 12 degrees from the hard palate line, as visualized on the lateral CT scout view. Eight- to 10-mm-thick sections are routinely obtained, but thinner slices with 1.5- to 3-mm thickness are necessary in evaluation of the orbits, pituitary gland, suprasellar and parasellar regions, and the CPA. In these situations, reformatted images in coronal and sagittal planes are useful. With the advent of the MR scanner, however, head CT is limited mainly to detect bleeding in acute stroke, acute trauma, calcification, and bony diseases.
Contrast Studies
Intravenous (IV) contrast medium is helpful to improve the detection and characterization of intracranial lesions. Enhanced CT scanning is not used in patients with acute trauma, patients with acute stroke to exclude a bleed, follow-up of trauma patients, patients with renal disease, or patients with a history of allergy to iodinated contrast medium. About 100 to 150 mL of a 350 mg I/mL iodinated contrast agent (adult dosage) should be administered for optimal study results.
MRI Technique
MRI parameters include repetition time (TR), echo time (TE), matrix size, field of view (FOV), slice thickness, and number of excitations (NEX). TR and TE are the only parameters that affect T1 and T2 weighting of the image. T1 images have a short TR of less than 1000 milliseconds and a short TE of less than 30 milliseconds. T2-weighted images have a long TR of more than 1500 milliseconds with dual echoes. The first echo of T2 images with shorter TE is called proton density, or a mixture of T1 and T2. The second echo of T2 images with a longer TR of more than 60 milliseconds represents real T2 images. T2-weighted images are most sensitive for detecting brain pathology, owing to the higher water content in most of the lesions.
Gadolinium compounds, which are paramagnetic IV contrast media for brain imaging, are especially helpful for identification of extraaxial tumors and intraaxial lesions associated with disrupted blood-brain barrier. The regular dose is about 0.1 mmol/kg of body weight, but in a case with equivocal findings or solitary metastasis, a dose of 0.2 to 0.3 mmol/kg of body weight is advisable. The same structures that enhance with IV use of contrast medium on CT scans enhance similarly on MRI scans.
Sectional Anatomy
Normal Axial CT and MRI Anatomy
On CT and MR scans the brain has been briefly viewed in infratentorial and supratentorial sections, as described below. CT scans are performed with a 15- to 20-degree angulation to the canthomeatal line at 8-mm increments. MRI scans are generally obtained parallel to the AC-PC line in the axial plane, with 6-mm slice thickness. Using the sagittal view the coronal sections are acquired parallel to the brainstem and the sagittal sections are obtained perpendicular to the axial section.
Infratentorial Sections
Below the fourth ventricle ( Fig. 8-3 ).
In this section, most of the structures in the anterior and middle fossa are components of the base of the skull and the orbits. In the middle fossa the foramen ovale and foramen spinosum can be visualized with a wide window setting on CT scans. Through these foramina pass the third branch of the fifth cranial nerve and the middle meningeal artery, respectively. The cerebellar hemispheres can be seen lateral to the medulla. On MRI studies, cranial nerves IX and X can be demonstrated at this level because they emerge from the postolivary sulcus. The posterior aspect of the cerebellar hemispheres is outlined by the inferior portion of the cisterna magna.

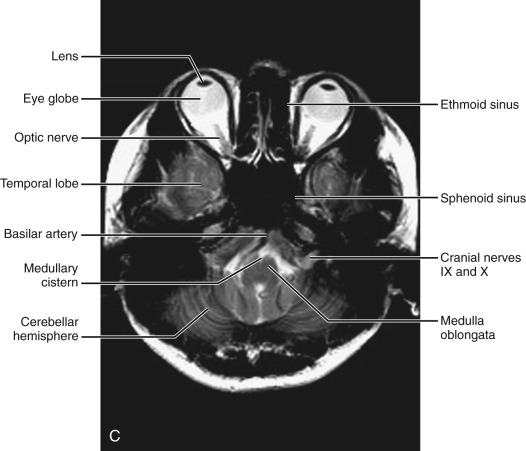
Fourth ventricular level ( Fig. 8-4 ).
The lower pons is seen in front of the fourth ventricle, connecting to the cerebellar hemispheres by the middle cerebellar peduncles. The pons is outlined by CSF, with the anterior and lateral pontine cisterns. Anterior to the pons appears the basilar artery, with signal void on regular MRIs. Posteriorly the fourth ventricle is outlined by the cerebellar nodulus in the midline and by the cerebellar hemispheres laterally. On MRI examinations the seventh and eighth cranial nerves can be seen at the CPA.


Above the fourth ventricle ( Fig. 8-5 ).
At the center, the midbrain appears outlined by the suprasellar cistern, ambient cistern, and quadrigeminal cistern. The suprasellar cistern contains the internal carotid artery, the anterior and middle cerebral arteries, the optic chiasm, the infundibulum, the mammillary bodies, and the top of the basilar artery. Lateral to the midbrain are the temporal lobes, where the uncus, amygdaloid nucleus, and hippocampal formation can be demonstrated. The temporal horn can be seen in the middle of the temporal lobe at this level. In the anterior fossa, the frontal lobes are separated from the temporal lobes by the sylvian fissure. Most inferior portions of the frontal lobe can be seen at this level. In the posterior fossa, the aqueduct of Sylvius is present behind the midbrain. The superior cerebellar surface is seen with separation of the two hemispheres by the superior vermis.



Supratentorial Sections
Third ventricular level ( Fig. 8-6 ).
The frontal lobes can be seen separated by the interhemispheric fissure medial to them. Superficially the sylvian fissures extend medially to separate the frontal lobe from the temporal lobe. Medial to the medial surface of the sylvian fissure, the insular cortex, external capsule, putamen, and globus pallidus can be demonstrated. The third ventricle in the midline is seen as a slitlike cavity surrounded by the thalamus laterally. Behind the third ventricle, the internal cerebral vein, the great vein of Galen, and the cerebellar vermis can be seen. Sometimes the pineal gland behind the third ventricle is seen at this level. Occasionally some calcification of the habenula occurs anterior to the pineal gland calcification. Within the lateral ventricle the most commonly calcified choroid plexus can be identified. Posteriorly the occipital lobes are visualized.



Low ventricular level ( Fig. 8-7 ).
The most superior aspects of the frontal lobes are visualized. The superior portion of the frontal horns appears outlined by the head of the caudate nuclei laterally. Anteriorly the frontal horns are shaped by indentation of the genu of the corpus callosum. The corpus callosum is seen separated from the cingulate gyrus by the cingulate sulcus. Posteriorly the occipital horns are indented by the splenium of the corpus callosum. Within the lateral ventricle the choroid plexus can be visualized on MRIs. Behind the splenium of the corpus callosum, the straight sinus is presented as signal void on regular MRIs.

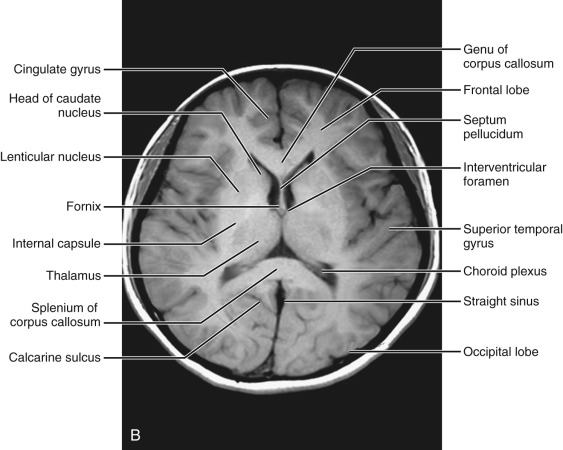

Midventricular level ( Fig. 8-8 ).
The superior extension of the sylvian fissure and the superior temporal gyrus are seen. The central sulcus separates the temporal lobe from the parietal lobe. The most superior aspect of the frontal horns is indented laterally by the most superior aspect of the caudate nuclei. The posterior medial aspect of the occipital horns is seen bound by the white matter fibers of the splenium of the corpus callosum. The posterior portion of the cingulate sulcus, separating the cingulate gyrus from the occipital lobe, can be seen connecting to the posterior interhemispheric fissure.

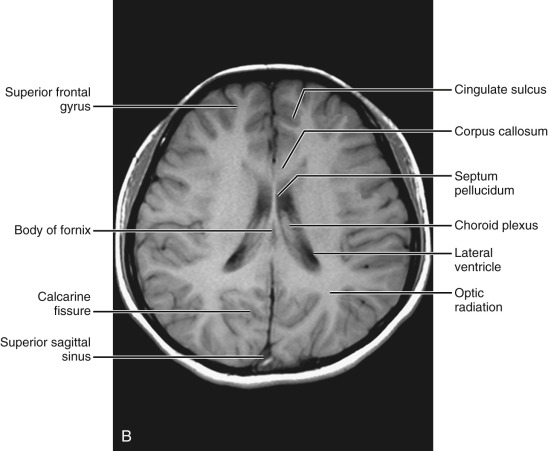

Above the ventricular level ( Fig. 8-9 ).
This section mainly includes the frontal lobe, parietal lobe, and a small portion of the occipital lobe. The superior frontal sulcus, separating the superior frontal gyrus from the middle frontal gyrus, is well delineated parallel to the interhemispheric fissure. Three parallel sulci including the precentral, central, and postcentral sulci are well visualized at this level. The central sulcus can be identified in the midportion of the scan. Precentral and postcentral sulci outline motor and sensory cortices. The interhemispheric fissure can be seen in its entire length, with the falx between. Lateral to the interhemispheric fissure the centrum semiovale, being massive white mater, is well documented.