Joti Jonathan Bhattacharya, Kirsten Forbes, Peter Zampakis, David J. Bowden, John M. Stevens
Overview of Anatomy, Pathology and Techniques; Aspects Related to Trauma
Overview of Anatomy, Pathology and Techniques
Modern imaging techniques depict the brain in ever more exquisite detail in all three orthogonal planes. Since neuroradiology forms an important part of radiology training and a substantial part of most radiologists’ daily work, familiarity with some of the intricacies of neuroanatomy becomes increasingly important; radiology remains in large part applied anatomy. The brain, at the macroscopic level, is a largely symmetric structure aiding the identification of abnormalities. Here we offer an overview of brain and vascular anatomy as shown on current imaging techniques, beginning with a brief summary of brain development. Imaging techniques for the brain and vasculature are then reviewed.
Anatomy of the Brain and Vascular System
Embryology
The brain derives from the rostral end of the embryonic neural tube, formed of neural ectoderm. The initially fairly uniform neural tube develops three swellings, the primordial cerebral vesicles (prosencephalic, mesencephalic and rhombencephalic), which subsequently give rise to five vesicles.1 At this stage it remains one cell thick, with a pseudostratified epithelium containing the neural stem cells. The cavity of the neural tube represents the future cerebral ventricles and the central canal of the spinal cord, ending anteriorly at the membrane of the lamina terminalis. Thus the anterior wall of the third ventricle (lamina terminalis) represents the rostral end of the neural tube (Fig. 60-1).
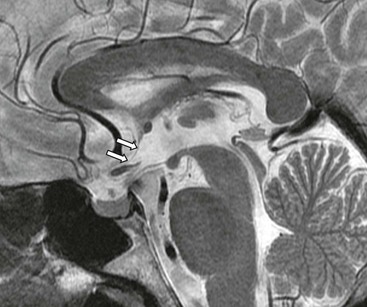
Bulges appearing on either side of the prosencephalic vesicle represent the developing telencephalic vesicles and, subsequently, cerebral hemispheres, and their opening, the future interventricular foramen (of Monro). With growth, the wall of the neural tube thickens and nutrition, which was initially by simple diffusion from the amniotic fluid to the neural plate, and after closure of the tube by diffusion from the surrounding primordial vascular plexus, is no longer sufficient. a depression appears in the roof of the developing third ventricle and adjacent cerebral hemispheres, invaginating a layer of ependyma and vascular pia mater, to form the choroid plexus (Fig. 60-2).
Thus the original function of the choroid plexus appears to be oxygenation and nutrition of the deep portions of the brain.2 Subsequently, penetrating vessels grow into the brain substance. With growth of the cerebral hemispheres, neuronal proliferation occurs in the periventricular zone, followed by migration of neurons along radially oriented glial cells to reach the pallial surface of the brain. This results in formation of the cerebral cortex, with its characteristic lamination. Over most of the hemispheric surface a six-layer neuronal structure can be identified: the isocortex (neocortex). Paul Broca traced the isocortex to its medial edge (Latin: limbus), thus identifying a medial limbic lobe (the hippocampus and associated structures) which was found to have a simpler three-layer structure, the allocortex. An intermediate band of cortex between the isocortex and allocortex can be discerned, termed the mesocortex. Other terminologies (archicortex, paliocortex, archipallium, etc.) are obsolete and better avoided. The developing cerebral hemispheres are initially separate, with the first crossing fibres developing in the lamina terminalis to form the anterior commissure. Superior to this, formation of the corpus callosum begins, progressing posteriorly. The surface of the hemispheres from an initially smooth (lissencephalic) appearance becomes progressively convoluted. The mesenchymal soft tissues overlying the brain, deriving mainly from the neural crest, differentiate into skull, dura mater and enveloping the brain the vascular meninx primitiva. Cavitation within the meninx primitiva separates the pia mater from arachnoid mater, producing the subarachnoid space, and modification of the primordial vascular plexus produces the surface vessels of the brain.
Cerebral Cortex, Lobar Anatomy and Deep Grey Matter Structures
If we consider neuroradiology begins with the first radiograph of a skull taken in 1895,3 we could imagine the disappointment of those pioneers as they realised that the brain was invisible to the ‘new radiation’. Indeed most of the history of neuroradiology has involved the quest for an image of the brain. This was achieved with the first CT images in 1972. The cerebral hemispheres can now be examined in superlative detail by MRI at 1.5 and 3 T, and high-field systems are offering the beginnings of MR microscopy, to probe details of the cerebral cortex.
The cerebral cortex itself consists of arrays of neurons (estimated to number 100 billion, each one communicating synaptically with many adjacent neurons in a system of astonishing complexity) which on Nissl staining appear to be arranged in layers.4,5 The cortex varies from 2 to 5 mm in thickness. Cortical thickness, though not its internal structure, is readily apparent on standard T1 IR and T2 sequences (Fig. 60-3).
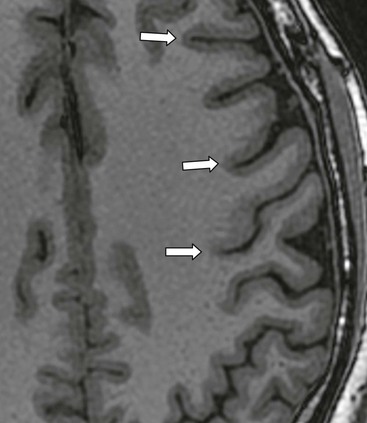
Isocortex, as described above, has six layers, allocortex has three, with mesocortex in between, all layers being numbered from superficial to deep. Probably, equal numbers of glial cells are present in the cortex, interacting metabolically with neurons and synapses as well as with the rich network of cortical capillaries. Precise functions of glial cells are not clear-cut, but they are certainly not mere supporting cells.6 The surface of the cortex is formed by a continuous layer of superficial astrocyte foot processes with associated basement membrane forming the glia limitans to which is applied the pia mater, with a potential subpial space intervening.
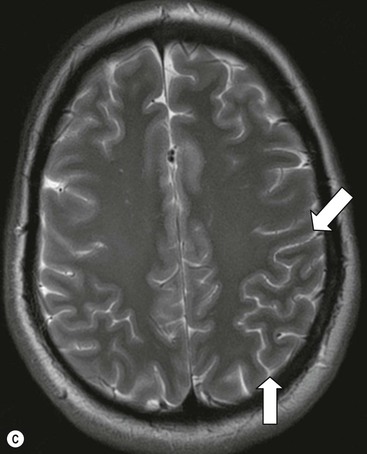
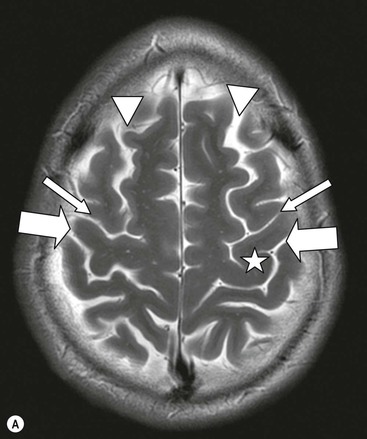
The cortical mantle over the surface of each hemisphere is folded into a series of elevated gyri separated by sulcal clefts. These form the basis of the separation into the lobes of the brain, which were originally named for the overlying skull bones. Gratiolet (in 1854) adopted this schema, adding precise boundaries which became widely adopted.7 The lobar terminology represents a convenient though arbitrary system (which has varied over time and between authors) and is largely devoid of ontogenic significance. Six lobes in each hemisphere are often described: frontal, parietal, occipital, temporal, insula and limbic. With familiarity, patterns emerge from the initially bewildering array of brain convolutions allowing quite accurate identification of the major subdivisions.8,9 The interhemispheric fissure and Sylvian (or lateral) fissure (Fig. 60-4) are immediately obvious.
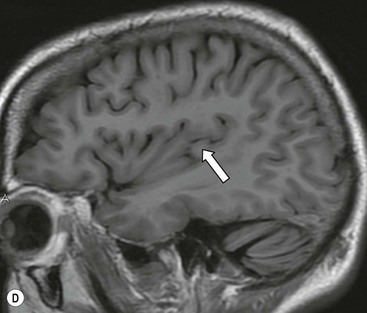
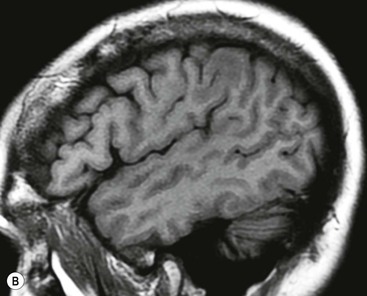
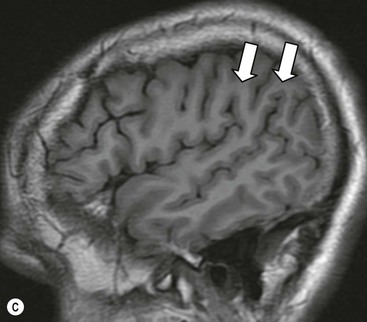
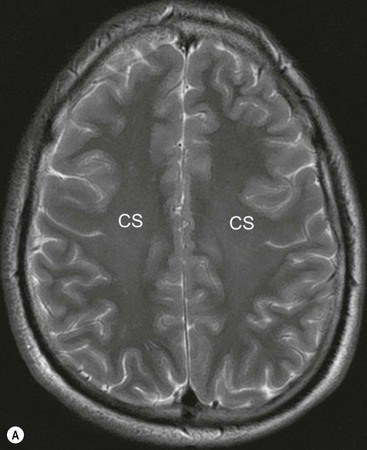
The central sulcus is the other main landmark of the hemisphere separating the precentral gyrus (motor) from the postcentral gyrus (sensory) and can usually be confidently identified on axial and sagittal images (Fig. 60-5). From these landmarks other sulci and gyri can be sequentially identified.8,10,11
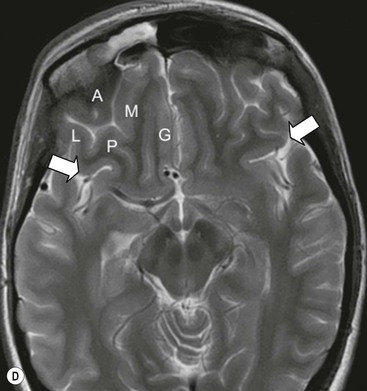
The deep grey matter structures principally comprise the basal ganglia, amygdala and thalamus and are well demonstrated by CT and MRI (Fig. 60-6). The basal ganglia are part of the extrapyramidal system including the caudate nucleus, globus pallidus, putamen, nucleus accumbens and substantia nigra. The globus pallidus and caudate are linked across the intervening internal capsule by a series of grey matter bridges giving a striated appearance, the origin of the term corpus striatum for this region.
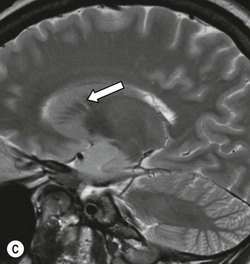
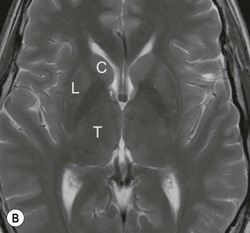
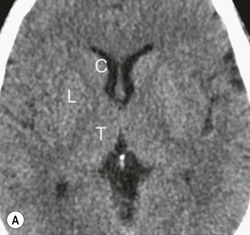
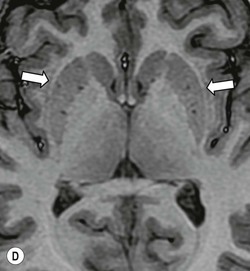
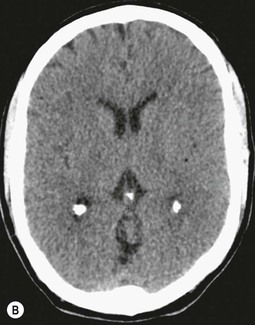
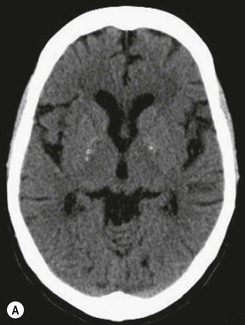
Beneath the internal capsule, these nuclei are linked by the nucleus accumbens. Physiological punctate calcification of the basal ganglia is commonly seen with ageing on CT images after the age of about 30 years.12,13 Iron deposition is also encountered in the basal ganglia, increasing with age from the second decade (Fig. 60-7).14 Similarly, calcification in the pineal gland is seen in about 40% of subjects by the age of 20 years (Fig. 60-8).15
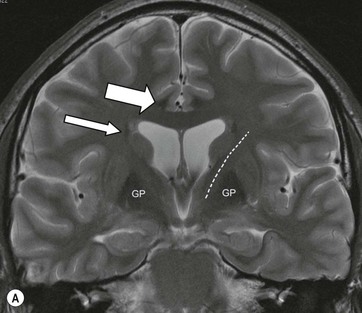
The thalami are paired large nuclear masses forming most of the lateral walls of the third ventricle, above and behind the hypothalamus. They often are in contact across the ventricle at the massa intermedia. The posterior border, or pulvinar, bulges convexly into the quadrigeminal cistern and overlies the medial (visual) and lateral (olfactory) geniculate bodies.
White Matter Centre
The anatomy of the white matter of the brain has generally received little attention in the imaging literature, which is surprising given the ubiquity of white matter diseases. The medullary core of the brain is formed of bundles of axons, supporting glial cells and penetrating blood vessels. Its whitish colour derives from the fatty myelin sheaths contributed by oligodendrocytes (in the periphery myelin sheaths are formed by Schwann cells). The lipid content accounts for the low density of white matter on CT images and for the characteristic high signal on T1 and low signal on T2 MRI sequences. The white matter is less metabolically active than grey matter and consequently receives a much smaller proportion of the brain’s blood supply. On axial anatomic or imaging sections the white matter core presents an oval aspect, Vieussens (eighteenth century) terming the component in each hemisphere, the centrum semiovale (Fig. 60-9).16 Long after Schwann (1839) established the cell theory, anatomists considered the white matter to be an amorphous continuum, which paradoxically, in imaging terms it has remained until recently. Tractography with MRI diffusion tensor imaging (DTI) can now reveal white matter bundles and their pathways in the living brain. Some of the larger tracts are apparent even on standard sequences. It is conventional in neuroradiology to describe lesions as lying in the subcortical (U-fibres immediately below the cortex), deep (white matter core) or periventricular (thin band of white matter adjacent to the ependyma) white matter.
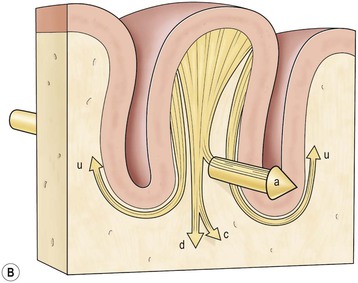
Axons entering or leaving the cortex can be classified into several types (Fig. 60-9). Thus subcortical U-fibres link one gyrus locally to adjacent gyri with a U-shaped band deep to the sixth layer of the cortex.
Ascending/descending tracts carry sensory information to the thalamus and cortex, or project motor fibres via the internal capsule to traverse the brainstem. For example, the corona radiata and internal capsule bearing the descending corticospinal tract, corticocerebellar tracts traverse the cerebellar peduncles and the ascending spinothalamic tracts.
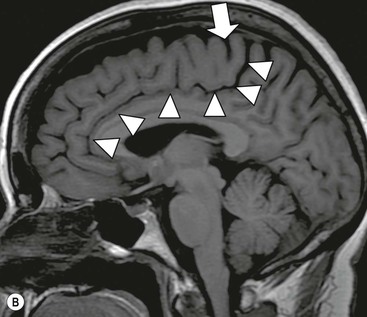
Association tracts link cortical areas in different lobes of the same hemisphere. The most prominent are the superior longitudinal fasciculus (SLF I, II and III) running in the white matter of the parietal lobe linking parieto-occipital and frontal lobes, the arcuate fasciculus, the extreme capsule, the fronto-occipital fasciculus (running with the subcallosal bundle of Muratoff, the combination being visible on standard T2 sequences MRI), the uncinate fasciculus linking anterior and mesial temporal structures with the frontobasal region, and the cingulum bundle, also visible on standard MRI (Fig. 60-7).
Commissural tracts are crossing fibres linking corresponding regions of opposite hemispheres, the largest of these being the corpus callosum, a mammalian innovation only absent in marsupials and monotremes. This dense bundle of fibres containing up to 190 million axons has a rostrum, genu, body and splenium. Anteriorly, its fibres fan out into the forceps minor and posteriorly from the larger splenium into the forceps major. Consisting of densely packed axons with relatively low metabolic requirements, it is less susceptible to ischaemic disease but is a typical site of demyelination, particularly on its ventricular surface. It also provides a common route of spread for aggressive neoplasms (butterfly glioma). The anterior commissure (AC), a dense bundle of axons, runs in the anterior wall of the third ventricle and is well demonstrated on sagittal and axial sections. The posterior commissure (PC), although more difficult to visualise directly on sagittal images, can be readily pinpointed since it is located at the point where the cerebral aqueduct opens into the third ventricle. The AC–PC line is a basic imaging plane for stereotaxic procedures and is thus easily drawn on sagittal images. There are several other smaller commissures, including the fornix (or hippocampal) commissure and the habenular commissure.
Limbic System, Hypothalamus and Pituitary Gland
Following the isocortical mantel over the hemisphere to its medial edges, the structures of the limbic system are encountered. These include the amygdala, hippocampus, parahippocampal gyrus, cingulate gyrus, subcallosal gyri and associated structures.
Limbic structures are associated with memory processing, emotional responses, fight-or-flight responses, aggression and sexual response: in summary, with activities contributing to preservation of the individual and the continuation of the species. These structures are demonstrated in detail by coronal and sagittal MRI. The limbic system is often rather misleadingly described as a phylogenetically ancient part of the brain: the hippocampus is unequivocally a mammalian innovation while the isocortex itself has equally ancient antecedents.
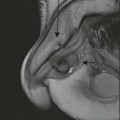
The core limbic structures are located in the medial temporal lobe readily amenable to high-resolution MRI. The amydala is the most anterior structure, separated from the hippocampal head by the uncal recess of the temporal horn (Fig. 60-10). The medial lying uncus (hook) has anterior amygdaloid and posterior hippocampal components.
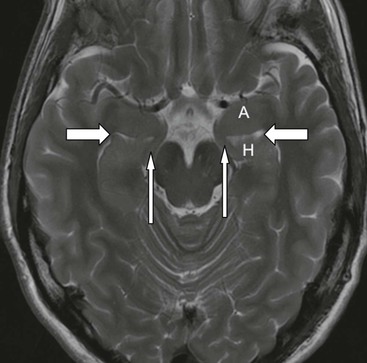
The hippocampal head, body and tail are well shown on coronal imaging, along with the parahippocampal gyrus (Fig. 60-11).
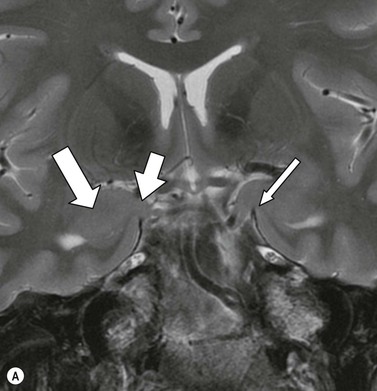
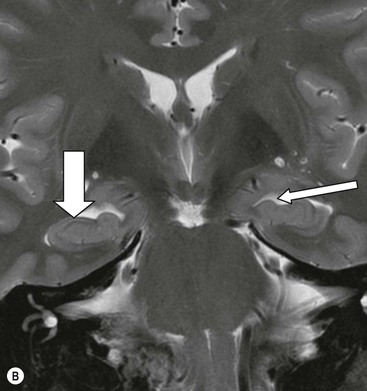
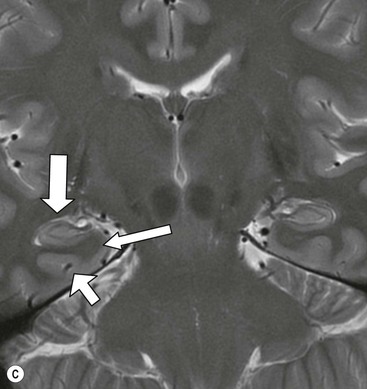
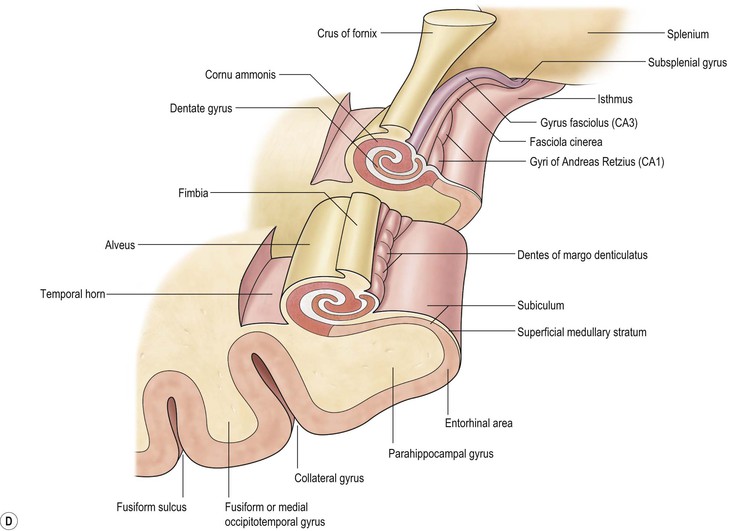
The white matter connections of the hippocampus via the fibria-fornix system are visualised on coronal and sagittal images. A thinned layer of hippocampal tissue, the indusium griseum, extends over the corpus callosum but is not visible on standard imaging.
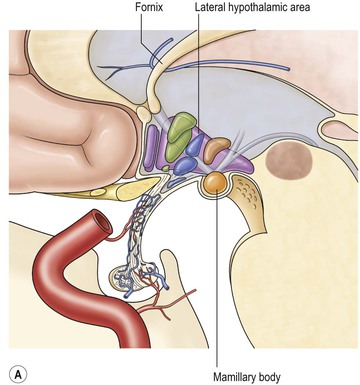
The hypothalamus forms the floor of the third ventricle and its side walls anteriorly following an oblique line inferiorly from the foramen of Monro to the midbrain aqueduct. It consists of a group of nuclei serving a number of autonomic, appetite-related and regulatory functions for the body as well as controlling and producing hormonal output from the pituitary gland. The hypothalamus is intimately linked to other limbic structures and might be considered the output for the limbic system.
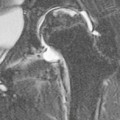
The pituitary infundibulum (or pituitary stalk), a hollow conical structure, extends inferiorly from the hypothalamus to the pituitary gland. The pituitary gland varies considerably in size, with sometimes only a thin rim of glandular tissue visible at the floor of the pituitary fossa. In young females, the gland may fill the fossa with a convex upper border. Anterior and posterior lobes can be distinguished on MRI, the posterior lobe often returning a high signal on T1-weighted images due to neurosecretory granules in the neurohypophysis. Both gland and stalk show strong contrast enhancement.
Ventricular System and Subarachnoid Space
The ventricular system, filled with cerebrospinal fluid, is the mature derivative of the cavity of the neural tube. Thus, the telencephalon contains the lateral ventricles; the diencephalon, the third ventricle; the midbrain, the cerebral aquaduct and the brainstem, the fourth venrticle (Fig. 60-12). The ventricles are lined by modified glial cells constituting the ependyma.
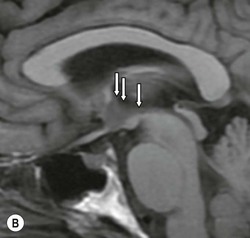
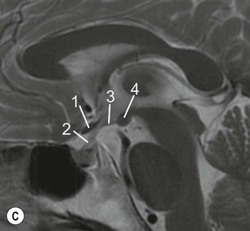
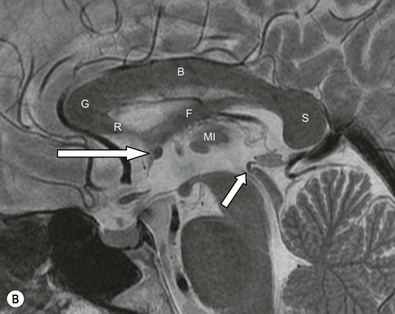
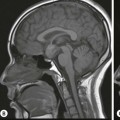
The lateral ventricles are divided into frontal horn, body, occipital (posterior) and temporal horns. The junction of the body, occipital and temporal horn is known as the trigone indented posteromedially by the calcar avis, the impression of the deep calcarine fissure. The sites of the original outpouchings of the telencephalic vesicles form the interventricular foramina of Monro linking the lateral ventricles with the anterosuperior aspect of the third ventricle. The third ventricle is a midline, slit-like cavity bordered laterally by the bulky grey matter of the thalami that frequently make contact with each other centrally as the massa intermedia. Antero-inferiorly and inferiorly lies the hypothalamus. A number of important structures are identified in the walls of the third ventricle on sagittal midline MRI slices (Figs. 60-7 and 60-13).
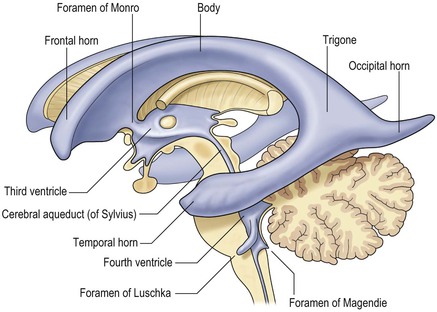
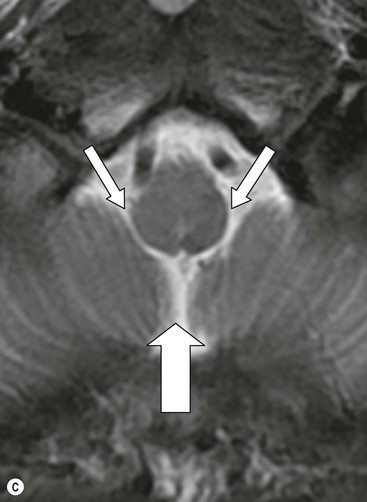
From the posteroinferior aspect of the third ventricle, the cerebral aqueduct (of Silvius) traverses the midbrain emerging into the rhomboid-shaped fourth ventricle (Fig. 60-14). The floor of the fourth ventricle is formed by the pons and medulla oblongata and it is roofed by the cerebellum, with its tented apex, the fastigium. There are two lateral apertures, the foramina of Luschka and a single posterior one, the foramen of Magendie allowing efflux of cerebrospinal fluid (CSF) into the subarachnoid space. At the inferior aspect of the fourth ventricle the embryonic continuation of the neural tube cavity into the central canal of the spinal cord is commonly obliterated in the adult.
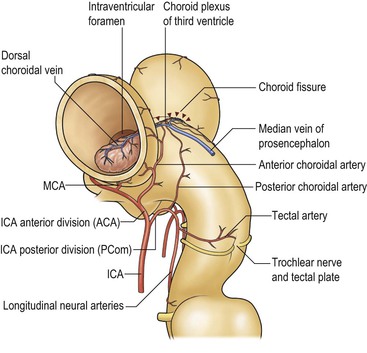
As described above, the cavity of the ventricular system is invaginated, from the choroid fissure by the choroid plexus, a vascular membrane with pial and ependymal layers. Anterior and posterior choroidal arteries enter this membrane. Choroid plexus is present in both lateral ventricles and extends through the interventricular foramina into the third ventricle forming the roof. In the fourth ventricle the choroid plexus extends laterally into both foramina of Luschka, often projecting into the subarachnoid space. Fourth ventricular choroid plexus is supplied by branches of the posterior inferior cerebellar artery.
CSF flows from the fourth ventricular foramina into the subarachnoid space (Fig. 60-15). Over the superior convexity of the brain, the subarachnoid space is thin. At the base conversely, because of irregularity of the inferior surface of the brain and skull, the spaces become widened in places which are known as subarachnoid cisterns, named for adjacent structures.
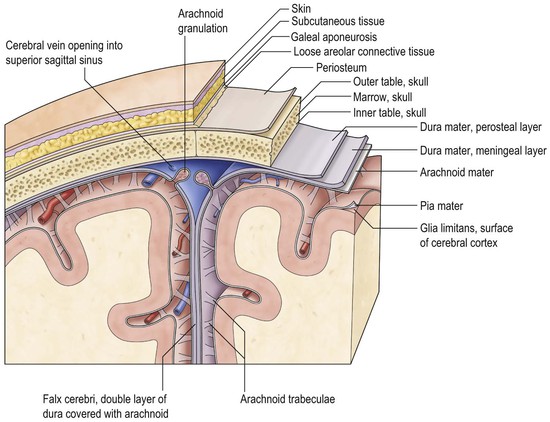
CSF is produced in part from the choroid plexuses, and in part from transudation of fluid from brain capillaries into the ventricles at a rate of approximately 500 mL per day. The total volume of CSF in the ventricles, intracranial and spinal subarachnoid space is about 150 mL. Thus, total CSF volume is turned over several times a day. Previous concepts that the majority of CSF is reabsorbed into the dural venous sinuses at the arachnoid granulations no longer appear tenable.17,18 Whether the granulations are involved at all in CSF reabsorption remains debatable and their major functions are unknown in adults as well as infants in whom they are not fully formed. The major routes of CSF reabsorption appear to be along the cranial nerve linings, large vessel adventitia and the cribriform plate into the lymphatic system, via the spinal, especially lumbar, epidural space as well as directly back into the capillaries and venules of the brain.
Cerebellum
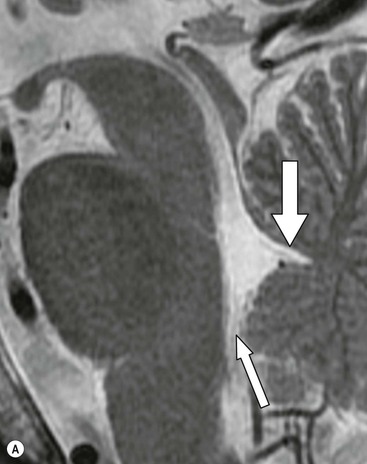
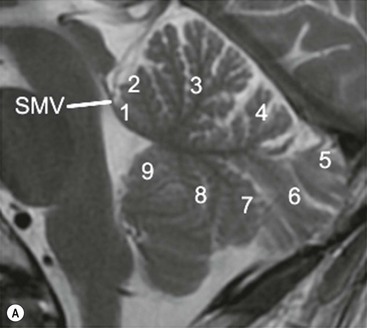
The cerebellum occupies the bulk of the posterior fossa, lying posterior to the fourth ventricle and brainstem to which it is linked by the white matter tracts of the paired superior, middle and inferior cerebellar peduncles. It is separated from the occipital lobes by the dural fold of the tentorium cerebelli. The cerebellum displays much finer folding than the cerebral hemispheres, the folds being termed folia. There are two cerebellar hemispheres joined by a midline portion, the cerebellar vermis, which is divided into a number of lobules readily identified on sagittal MRI (Fig. 60-16). Cerebellar cortex overlies the cerebellar white matter. Within the white matter on each side are the cluster of paired deep cerebellar nuclei, the largest of which are the two dentate nuclei. The principal functions of the cerebellum involve the coordination of skilled voluntary movements and muscle tone, each cerebellar hemisphere serving this function for the same side of the body.
Brainstem
The brainstem is usually considered to include the midbrain, pons and medulla oblongata and extends from the posterior commissure at the opening into the third ventricle to the pyramidal decussation at the cervicomedullary junction. This may be divided into the hindbrain or rhombencephalon, comprising the medulla oblongata and pons (as well as the cerebellum which develops from them) and the mesencephalon or midbrain.
The midbrain (Fig. 60-17) consists of the anteriorly lying cerebral peduncles containing the descending and ascending tracts. These are separated by the grey matter nuclei of the substantia nigra from the midbrain tegmentum. The tegmentum extends posteriorly to the cerebral aqueduct (of Silvius). Posterior to the aqueduct lies the tectal (or quadrigeminal) plate with its superior and inferior colliculi. Cranial nerve IV arises here also, the only cranial nerve to arise from the dorsal surface of the brainstem. Grey matter structures within the midbrain include the substantia nigra and red nuclei, readily identified on MRI, as well as the upper cranial nerve nuclei. The medial forebrain bundle bringing limbic fibres from the septal area and hypothalamus ends in the reticular formation, a loose array of neurons, part of the limbic midbrain.
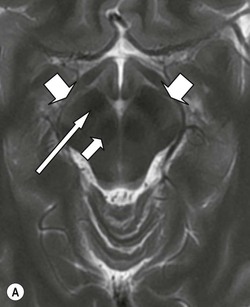
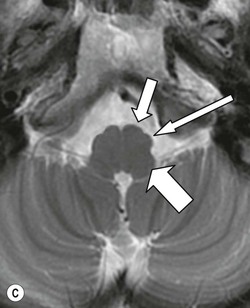
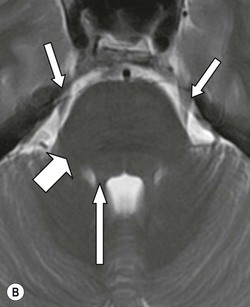
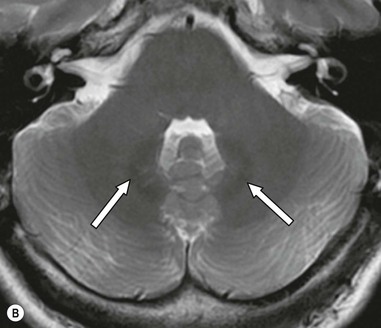
The pons (Fig. 60-17) comprises a bulbous convexity anteriorly, the basis pontis, containing masses of transversely oriented fibres which enter the large middle cerebellar peduncles on each side. Amongst these fibres are the dispersed bundles of the corticospinal tracts which separate as they leave the midbrain and reform as they enter the pyramids of the medulla. The posterior part of the pons, the pontine tegmentum, forms the floor of the upper part of the fourth ventricle, and contains cranial nerve nuclei (V, VI, VII, VIII).
The medulla oblongata (Fig. 60-17) consists of an inferior closed portion, with its central canal extending into the spinal cord, and a superior open portion related to the inferior portion of the fourth ventricle. The closed part of the medulla extends from the C1 spinal roots to the obex at the lower margin of the fourth ventricle. The ventral surface of the medulla is marked by two bulges on each side: medially lie the pyramids and laterally, the olives. The medulla transmits ascending sensory tracts posteriorly and descending motor tracts anteriorly both of which cross the midline, or decussate within the closed medulla (sensory decussation lying slightly higher). The lower cranial nerve nuclei lie within the medulla but are not visible with imaging. In cross-section the medulla oblongata is, however, well demonstrated on MRI.
Cerebral Vasculature
The internal carotid arteries (ICA) supply the anterior cerebral circulation while the vertebral forming the basilar artery supply the posterior circulation (Fig. 60-18), the two meeting at the circle of Willis. The external carotid arteries (ECA) supply most extracranial head and neck structures (except the orbits) and make an important contribution to the supply of the meninges. There are numerous anastomoses between the external carotid arteries and the anterior and posterior circulation.
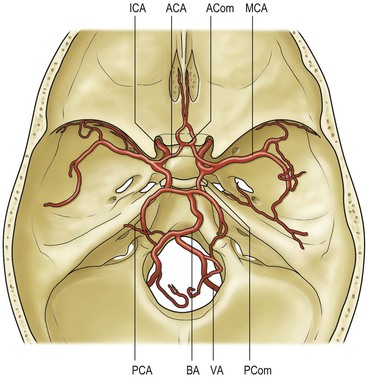
The aortic arch gives rise to three main branches, brachiocephalic, left common carotid and left subclavian arteries. The common carotid arteries run within the carotid sheath, lateral to the vertebral column, and bifurcate usually at the fourth cervical vertebrae into external and internal carotid arteries.
Anterior Circulation
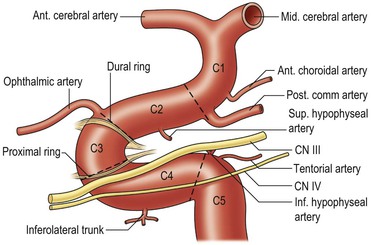
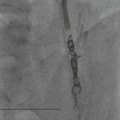
The internal carotid artery can be divided into a number of segments C1–C5. The cavernous segment (Fig. 60-19) gives branches to dura, pituitary gland and cranial nerves before its first major branch, the ophthalmic artery.19 The tentorial and inferior hypophyseal vessels may arise as a meningohypophyseal trunk. The inferolateral trunk supplies adjacent cranial nerves and anastomoses with the ECA. After leaving the cavernous sinus, it pierces the dura and enters the subarachnoid space at the level of the anterior clinoid process (Fig. 60-20). The posterior communicating artery is of very variable size, sometimes occurring as a fetal-type posterior cerebral artery. The anterior choroidal artery supplies the posterior limb of internal capsule, cerebral peduncle and optic tract, medial temporal lobe and choroid plexus. The supraclinoid segment of the ICA divides into the anterior (ACA) and middle cerebral arteries (MCA).
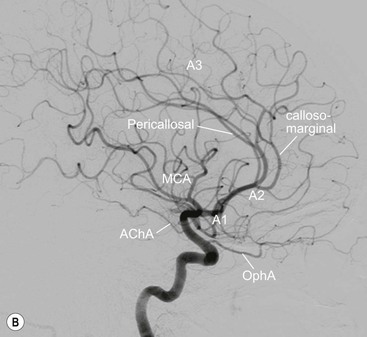
The anterior cerebral artery20,21 is divided into three anatomical segments: horizontal or precommunicating segment (A1), vertical or postcommunicating segment (A2) and distal ACA including cortical branches (A3). The A1 segment is joined to the contralateral A1 segment by the anterior communicating artery. The A1 segment gives rise to perforating branches, the medial lenticulostriate arteries. The recurrent artery of Heubner is the largest of the perforating branches doubling back on the parent artery, arising from the A1 or A2 segment. One or other A1 segment may be hypoplastic, or the A2 segments may fuse to give a midline azygous anterior cerebral artery. Cortical branches of the ACA supply the medial aspect of the cerebral hemisphere.
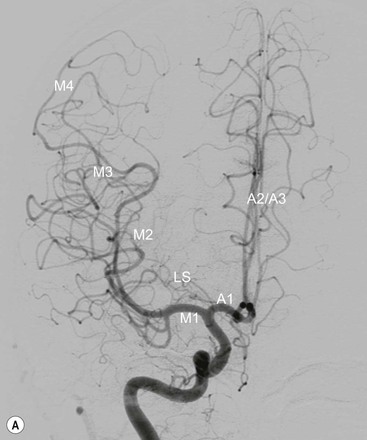
The middle cerebral artery22 is divided into four anatomical segments: horizontal segment (M1), insular segment (M2), opercular segment (M3) and cortical branches (M4 segments). Medial and lateral lenticulostriate arteries are perforating branches that arise from the M1 segment supplying the basal ganglia and capsular regions. The M1 segment divides as a bifurcation or trifurcation. Cortical branches supply the lateral surface of the cerebral hemispheres.
Posterior Circulation
The vertebral arteries usually arise as the first branches of the subclavian arteries (V1), then entering the foramen transversarium of the sixth cervical vertebra. They run upwards through each vertebra (V2) before arching around the anterior arch and behind the lateral mass of the atlas (V3) to pierce the dura mater and enter the sub-arachnoid space at the level of the foramen magnum (V4), fusing with their fellow in front of the lower pons, to form the basilar artery (Fig. 60-21). The left vertebral artery is commonly larger. The vertebral arteries give muscular branches, which anastomose with the ascending pharyngeal and occipital arteries, posterior meningeal branches, as well as supplying the cervical spinal cord. After entering the cranial cavity, each vertebral artery gives off a posterior inferior cerebellar artery (PICA) supplying the brainstem, inferior cerebellar hemisphere, vermis and the choroid plexus.
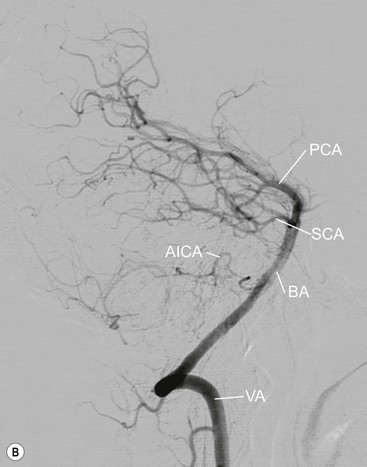
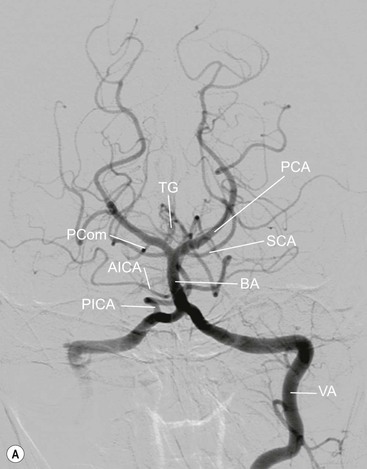
The basilar artery runs superiorly on the anterior surface of the pons giving off anterior inferior cerebellar (AICA), superior cerebellar (SCA) and posterior cerebral arteries on both sides as well as perforating branches. It terminates just above the tip of the dorsum sellae.23
The AICAs enter the cerebellopontine angle, and supply the surrounding structures; their branches include the internal auditory arteries, to the nerves in the internal auditory meatus. The cerebellar branches anastomose with those of the PICA. The SCAs arise several millimetres below the posterior cerebral arteries from which they are separated by the tentorium cerebelli. They pass around the brainstem to fan out over the superior surface of the cerebellar hemispheres.
Related posts:
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


