Fig. 1
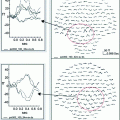
MEG source estimates. Dynamic Statistical Parametric Maps (dSPM) averaged in sequential 50 ms time bins from 200 to 500 ms for the good reader group (top) and the poor reader group (bottom). The group-averaged dSPM for the two subtraction contrasts (phonologically similar (PS) and phonologically dissimilar (PD)) are shown on the reconstructed cortical surfaces of one child. The lateral surfaces have been inflated for better visualization of activation within the sulci (dark gray) as well as the gyri (light gray). Both left and right hemispheres are shown for all time points. Note, delayed and less lateralized response to PS compared to PD condition in poor readers (Mody et al. 2008)
Salmelin et al. (1996, 2000) found that good readers show a strong response to reading words relative to nonwords 150–200 ms post-stimulus in OT area, a region known to be sensitive to letter-strings. Dyslexic readers did not display this early response, which may account for the subsequently weaker and delayed activation in semantic processing in temporo-parietal areas (Helenius et al. 1999). These studies suggest that a disruption in the grapheme-to-phoneme translation interface can have consequences for comprehension. In fact, the temporal profiles of the activity in language-implicated frontal, temporal and parietal areas reflect near-simultaneous peaks in children with reading difficulties compared to the distinct temporal progression of activity associated with component reading processes in these areas, in a typical reader (Rezaie et al. 2011). It is important to note that phonological processing skills continue to develop into mid childhood, as evidenced by changes in underlying speech perception skills. Parviainen et al. used MEG to record neural responses to speech and nonspeech sounds in 7–8 year olds and adults (Parviainen et al. 2011). Whereas both groups activated the superior temporal cortex, the differentiation between the cortical response to the two types of sounds was later (250 ms after sound onset), more prolonged and evident in both hemispheres in children; adults showed a corresponding effect though earlier, at 100 ms post-stimulus, and specifically in the left hemisphere which was linked to superior reading skills. Findings of less lateralized responses to auditory stimuli in children with dyslexia may suggest a problem in balance of auditory functions between the two hemispheres (Johnson et al. 2013).
In a recent study, Han et al. (2012) used MEG to investigate neural oscillatory activity associated with auditory sentence processing in reading impaired adolescents and age and IQ-matched controls. Insofar as auditory comprehension involves the integration of sensory, cognitive, and linguistic processes to arrive at the meaning of a word or sentence, neural synchrony may be a promising target mechanism for the study of higher-level language. The authors used an N400 task from a previous study with younger dyslexic readers (Mody et al. 2008) to examine top-down and bottom-up interactions in integrating sentence-terminal phonologically similar (PS) versus dissimilar (PD) incongruent words into preceding context. Similar to the results from the previous study, older good and poor readers also differed in their brain activation as a function of the degree of phonological contrast but in the time range of semantic processing (400–600 ms). Specifically, the functional coupling between auditory and superior temporal cortices in the right hemisphere as measured by PLV, was greater in the PS than PD condition for good readers. The results were interpreted as reflecting good readers’ superior coding of subtle phonetic differences between perceptually confusable terminal words in a semantically conflicting context. In contrast, for poor readers, who were selected on the basis of impaired decoding abilities, the PLV was lower in the PS than in the PD condition. This may relate to the group’s weak speech perception of subtle phonological differences exacerbated under conflicting semantic cues. That the group difference in phase-locking was significant in the gamma band is in keeping with existing studies that have indicated that gamma-band oscillations may play an important role in spoken language comprehension (Shahin et al. 2009; Obleser and Kotz 2011).
According to a recent study, poor readers’ difficulties may arise in part because of impaired phase locking to the slower modulation in the speech signal (<10 Hz), which could affect syllable segmentation abilities key to processing phonological aspects of speech (Hamalainen et al. 2012). To the extent that the responses patterns of the dyslexic and unimpaired readers in the study by Han et al. (2012) was modulated by the degree of phonological contrast, the results appear to be consistent with a phonological rather than an auditory temporal account of the disorder. The findings are also in keeping with the lack of a consistent auditory brainstem response to complex speech-like sounds and not to simple clicks in impaired readers, seen in EEG (Hornickel and Kraus 2013), which was taken as evidence of poor neurophysiological coding and may be viewed as further support of weak and underspecified speech sound representations in dyslexia.
4.2 Autism
Unlike dyslexia, which is primarily a written language disorder, individuals with autism have spoken language problems (Hudry et al. 2010). Early on the problem takes the form of a lack of orientation to speech and to their name when called (Osterling et al. 2002; Nadig et al. 2007). These behaviors have been found to be predictive of broader receptive language problems. However, the apparent difficulty with auditory comprehension may also be reflective of difficulties with testing this population due to a lack of social motivation.
Despite the absorption of language deficits under the broad umbrella of social communication deficits in the new DSM-5, language impairment remains a striking feature of ASD. In general, semantic deficits are among the few consistent language findings in autism (Rutter et al. 1992; Tager-Flusberg and Joseph 2003; Vogindroukas et al. 2003). While the number of functional neuroimaging studies of speech and language in autism has been limited, recent auditory event-related potential studies have begun to yield some important insights into the nature of the language impairment in ASD. Measures like the MMN response, which is larger for between- than within-category phonetic differences, has the potential to reveal abnormalities in auditory perception which have been implicated in autism and dyslexia. Čeponiene et al. (2003) used EEG with an oddball paradigm to examine the MMN for speech (viz., vowels) and non-speech stimuli (viz., simple tones, complex tones) each with a corresponding frequency deviant (created by raising all the frequencies of the original token by 10 %) in high-functioning children with autism and age-matched controls. Whereas the authors found that there was no difference in the MMN between the groups for all three stimulus types, revealing intact auditory processing in ASD, some studies have yielded delayed MMN responses to speech and non-speech in autism (Oram Cardy et al. 2005; Roberts et al. 2011), similar to that observed in children with specific language impairment (Roberts et al. 2012), and indicative of language impairment in autism. In these latter studies, the speech stimuli consisted of two different vowels, /a/and/u/; Čeponieni et al. however, used a standard vowel paired with an acoustic deviant that was not phonologically different, which might explain the different results. Nevertheless, it is interesting, then, that the ASD participants while showing a normal MMN response, lacked the P3a response (a neural index of involuntary orienting to a novel or salient stimulus) in the speech condition, though not in the tone conditions. In keeping with the literature, the ASD participants did not orient to speech which may account for difficulties in auditory comprehension and an apparent preoccupation with their own world.
In older individuals on the spectrum, abnormal functional organization in fronto-temporal areas has been consistently observed in fMRI tasks of spoken and written language comprehension. These semantic tasks have included priming, categorization, fluency, and sentence processing. Activation is typically reduced in the left hemisphere (Just et al. 2004; Harris et al. 2006; Gaffrey et al. 2007) and increased responsiveness is evident in the right hemisphere (Wang et al. 2006; Knaus et al. 2008; Mason et al. 2008). Studies of reading in high-functioning autism suggest that individuals on the spectrum do not take advantage of contextual cues (Happe 1996). Using a sentence anomaly paradigm, Braeutigam et al. (2008) found that the neuromagnetic N400 response following incongruous words was weaker over left temporal cortices in individuals with autism. The incongruent sentence terminal words also elicited long-lasting gamma oscillations above 40 Hz in the ASD group but not in the controls. While the latter finding is hard to interpret, the results point to atypical semantic processing in ASD. Interestingly, access to semantics via pictures as well as picture naming appears to be less affected in autism (Kamio and Toichi 2000; Walenski et al. 2008; Sahyoun et al. 2010b) supporting an apparent dichotomy between visuo-spatial and linguistic abilities in autism (Tager-Flusberg and Joseph 2003; Behrmann et al. 2006). In separate fMRI and DTI studies, Sahyoun et al. used a three-condition pictorial problem solving task designed to vary the linguistic processing demands across the conditions (Sahyoun et al. 2010a, b). The authors found no difference in behavioral performance between high-functioning children with autism (HFA) and age- and IQ-matched controls regardless of linguistic demands; however, the control group relied more on frontal and temporal language areas whereas the HFA group activated occipito-parietal and ventral temporal areas. Taken together with findings of reduced white matter integrity of the connections between inferior frontal and middle/ventral temporal areas, the results appear to support HFAs’ preference for visuo-spatial strategies in the face of linguistic weaknesses. They also provide connectivity patterns that lend themselves to further fine-grained probing by MEG through use of language tasks that implicate temporal coupling of the same brain areas.
To date, studies of language processing with MEG in autism are limited. While there have been several studies of basic auditory processing in autism, few have examined higher level language processing. The auditory studies involving speech have, for the most part, looked at sound discrimination early in the auditory pathways using the MMN. The results have been mixed, with some reporting abnormal MMN responses (Jansson-Verkasalo et al. 2003; Roberts et al. 2011), yet others finding no abnormalities (Kemner et al. 1995; Čeponiene et al. 2003). Additionally troublesome is the lack of reliability in determining an MMN response, which raises questions as to its usefulness in clinical screening (Kurtzberg et al. 1995). Recent findings of abnormalities in gamma-band oscillatory responses during a continuous word recognition task in parents of children with ASD suggest a possible role for neural oscillatory responses in the search for a heritable neurophysiological biomarker of ASD (McFadden et al. 2012).
5 Future Directions
Findings from a number of behavioral and MEG studies appear to be converging on core abnormalities in dyslexia and autism. Evoked responses and neural synchrony measures in basic and intervention research appear to support a phonological core deficit in developmental reading disability. In later years, the problem of accuracy, frequently gives way to one of fluency, with “slow but accurate” reading frequently being the hallmark of the remediated adult dyslexic reader (Shaywitz et al. 2003). EEG and MEG with their superior temporal resolution are ideally suited to temporally unravel the interactions between top-down and bottom-up processes in reading. Such an approach could be tremendously powerful in a comparison, for example, of reading comprehension in dyslexia versus autism, insofar as a large number of individuals on the spectrum tend to be hyperlexic (i.e., precocious decoders). The findings from such contrasting disorders would move us closer to understanding the neurocognitive architecture of language, as well as the nature and locus of speech and language deficits critical to solving the disconnection puzzle that appears to be characteristic of neurodevelopmental disorders like autism and dyslexia (Mody et al. 2013). Finally, the development of passive task paradigms that reliably capture language processing will go a long way in extending the use of MEG and EEG to the study of minimally verbal ASD, a much needed area of research.
References
Abell F, Krams M, Ashburner J, Passingham R, Friston K, Frackowiak R, Happe F, Frith C, Frith U (1999) The neuroanatomy of autism: a voxel-based whole brain analysis of structural scans. NeuroReport 10:1647–1651
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


