Fig. 1
First five bars of the first movement of sonata no. 14 (“Moonlight”) by Ludwig van Beethoven (Op. 27, No. 2). The excerpt shows the characteristic triplet pattern in the right hand and the deviations from the pattern within the first bars. (Music typeset and published in the public domain by Chris Sawer, www.mutopia-project.org)
Unexpected deviations from a regular sound pattern are a powerful tool in musical composition, but they are also common in everyday life and often convey important information. Consider a crack of a branch standing out in the acoustic environment of a nightly scene in the woods that might indicate the approach of a predator, or a sudden change in the ongoing, regular noise of a car engine that might indicate an engine malfunction. Research on the encoding of sounds at the level of the auditory cortices has shown that we automatically encode various types of regularities in our acoustic environment, and that neuronal networks automatically detect deviations from regularities (Kujala et al. 2007; Näätänen et al. 2007). The mismatch negativity (MMN or MMNm) is a component of auditory evoked responses in EEG and MEG, respectively, that is sensitive to such deviant sounds in a sequence of regular sounds and to violations of expectancies that have been created by the preceding acoustic context. It occurs approximately 150–250 ms after the onset of the deviant sound, with a similar field distribution as the N1m component (auditory evoked response at approx. 100 ms latency), and it is superimposed on the auditory evoked field elicited by the sound. The MMN is a widely used tool in basic and clinical neuroscience that is also useful for research on music processing (Kujala et al. 2007; Näätänen et al. 2007).
Several MEG studies have looked at how we process short melodies that consist of a few consecutive tones. By using the MMN as an indicator, we can investigate which types of sound changes are detected at the level of the auditory cortices. We can also deduce what information from the stimulus is encoded, because the violation of a rule or regularity can only be detected if the underlying rule that is being broken was encoded in the first place. Classic MMN studies employ so-called oddball paradigms, in which one sound is presented with high probability and another sound is presented more rarely (Näätänen and Alho 1995). This deviant sound can be a tone of different pitch, duration, timbre, loudness or another physical parameter. If the difference between the standard and deviant sounds is large enough, a MMN response is observed in the magnetic field in response to the deviant event. For research on music, this basic approach can be adapted to more complex stimuli such as tone sequences. Here, the standard that is being repeated is not a single tone, but a short melody, and the deviant melody differs in some aspect. Both unfamiliar tone sequences (Boh et al. 2011; Fujioka et al. 2004) and familiar melodies (Hashimoto et al. 2000; Yasui et al. 2009) have been used in melody mismatch studies, and these studies have shed light on the complexity of material that can be stored, on the types of deviations that can be detected and on the capacity of the memory store underlying the deviance detection mechanism.
Fujioka et al. (2004) investigated the processing of melodies under passive listening conditions while participants watched a soundless movie. They presented simple five note melodies that varied in key from trial to trial. Deviant melodies either contained notes that changed the contour of the melody, or that differed only in the size of the musical interval between the tones, without a contour change. They found that while musicians showed clear MMN responses to both types of changes, nonmusicians only showed very weak responses. This was not due to a generally lower MMN response though, since both groups showed a similar MMN response in a simple oddball paradigm, indicating that the complexity of the material was a challenge for the automatic encoding in the nonmusicians.
In order to estimate the capacity of the auditory short-term memory storage underlying the mismatch detection mechanism for unfamiliar material in both musicians and nonmusicians, we designed a melody oddball study using unfamiliar melodies (Boh et al. 2011). In several conditions, we presented melodies of different lengths ranging from 4 to 8 tones. Deviant tones were equally likely to occur at all positions of the melody, and therefore the whole melody had to be encoded and stored in order to detect the deviants. We found that under such challenging circumstances the average capacity of auditory memory underlying the mismatch detection for musically untrained people is approximately 4 tones. For musicians however, this capacity was at least 8 tones, since they showed significant MMN responses to deviants even in the longest melodies (Fig. 2). The improved detection of melody deviants in complex, unfamiliar musical material is most probably due to their long-term musical training.
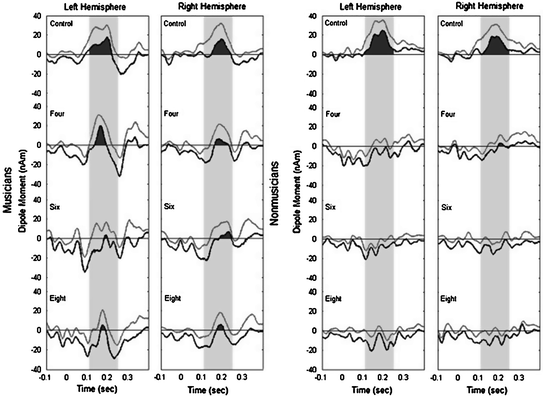
Fig. 2
MMN responses to pitch deviants in melodies of different lengths in musicians and nonmusicians in an MEG study by Boh et al. (2011). The grey traces show the group averaged source waveforms in both hemispheres, and the black traces show the lower confidence intervals as estimated from a bootstrap procedure. Nonmusicians show significant MMN responses only in the control condition (standard oddball paradigm) and in response to deviants in the four tone melody, whereas musicians show significant responses in all conditions. This result suggests that long-term musical training might lead to an increase of the capacity of auditory short-term memory for complex auditory patterns (Boh et al. 2011)
Even people without musical training automatically encode repeatedly presented short familiar melodies without having to attend to the stimuli. Hashimoto et al. (2000) presented participants without musical background with familiar melodies. Half of the melodies contained an incorrect note that was out of key of the original melody. These notes elicited an early mismatch response that was recorded with a limited number of channels over right auditory areas. Yasui et al. (2009) also used familiar melodies as stimuli. In a series of three experiments they differentiated the processing of lyrics and melody in familiar songs, and elucidated the role of long-term memory in the detection of unexpected events. They were able to show a respective right- and left-lateralization for melody and lyrics deviants in mismatch responses that were similar, but not identical in latency and source location compared to pitch MMN. Additionally, they asked participants to memorize new melodies and again they found the same mismatch responses to unexpected tones in these newly memorized melodies as in the melodies that were familiar from childhood. These findings indicate that not only short-term knowledge about regularities gained from the most recent auditory context but also long-term familiarity with melodies can be the basis for automatic deviance detection.
Some investigators have also looked at processing of musical chords and chord progressions using MEG. In most musical cultures and styles musical harmonies are organized in a musical syntax. People acquire knowledge about these rules via passive exposure in everyday life. This is evident from findings indicating that no special musical training is required to automatically detect untypical chords (Maess et al. 2001) and to distinguish major from minor chords (Brattico et al. 2009). Brattico et al. (2009) investigated the processing of chords in an MMN paradigm, where general musical context, in terms of mostly consonant major Western chords, was presented, but without a particular music-syntactic sequence or order of the chords. In this context, both musically trained and untrained participants showed a MMN response to rarely presented minor chords, indicating that even without a context of musically meaningful chord progressions such distinctions are automatically made. Maess et al. (2001) presented their subjects sequences of chords that followed Western musical rules of harmony. Infrequently, they presented chords that were consonant but did not quite fit at this position of the chord sequence. Such chords elicited the magnetic equivalent of another early response to unexpected sounds, the early right anterior negativity (ERAN). Importantly, the sources of the mERAN were estimated to be located in Broca’s area and its right homologue, and thus differed from typical sources of the MMN in the temporal lobes. This indicates that the syntactic processing and the detection of deviations in a musically meaningful harmonic context relies—at least partly—on a different network than the detection of deviant tones in unfamiliar tone sequences.
Another MEG study using the MMN as a marker for auditory processing investigated musical versus phonological processing (Tervaniemi et al. 1999). MEG provided the possibility to disentangle the respective contributions of the two hemispheres for processing of the two different stimulus types. Tervaniemi et al. (1999) showed that in the right hemisphere, the MMN to an unexpected chord was stronger than the MMN to a phoneme change, whereas there was no difference between the MMN amplitudes in the two stimulus categories in the left hemisphere. Furthermore, in an analysis of the source locations of the corresponding equivalent current dipoles, they found that the MMN sources of the two stimulus categories were distinct, indicating a specific neuronal network for processing of musical versus speech stimuli.
Apart from the spectral and pitch aspect of music that is most evident in the instrumental timbres and in the melodic and harmonic structure of music, another crucial component of music is the rhythmic and metrical structure of a piece. Vuust et al. (2005) investigated rhythm and meter processing in jazz musicians and in persons without musical background. They found that jazz musicians are especially sensitive to subtle deviations in a rhythmic sequence of percussion sounds that did not change the meter underlying the rhythm, whereas nonmusicians were only able to detect the more obvious violation of the meter. Interestingly, the deviance detection was right-lateralized in nonmusicians, but left-lateralized in musicians, again most probably due to the long-term musical training that resulted in changes in functional brain organization.
In summary, these studies demonstrate automatic detection mechanisms on the level of the auditory cortices that respond to unexpected or deviant auditory input. These mechanisms might also be related to some of the effects that make music interesting and beautiful. Although this has not been shown directly, the early and automatic detection of expectancy violations might contribute to emotional responses to unexpected changes in melody, harmony, dynamics or orchestration in music. However, more work is required to fully understand how the processes that detect regularities and expectancy violations have been shaped by nonmusical survival requirements, how music might rely on these evolutionary old mechanisms, and how it interacts with other systems such as language and cognitive systems that model regularities in our environment as a basis for expectancies and planning that are needed for adaptive behavior.
3 Higher-Order Cognition: Music in the Mind
In the beginning of the previous section we invited you to imagine some familiar music as an illustration of repeated tone patterns in music. If you know the piece or if you were able to sight-read it from the score, then you probably had an auditory impression that was a different experience from actual listening to music, but that nevertheless contained important aspects of the musical piece, such as the melody and accompanying chords, the piano timbre, the tempo and the dynamics, maybe even characteristic details of a particular recording that you remember.
Several MEG studies investigated what happens in the brain during higher-order musical cognition such as imagining and remembering music. In one study on auditory imagery (Herholz et al. 2008
 Decoding and Brain Machine Interfaces Based on Electromagnetic Oscillatory Activities: A Challenge for MEG
Neuroscience: A New Frontier for Magnetoencephalography?
Decoding and Brain Machine Interfaces Based on Electromagnetic Oscillatory Activities: A Challenge for MEG
Neuroscience: A New Frontier for Magnetoencephalography?
 Magnetoencephalography (fMEG)
Magnetoencephalography (fMEG)
 and Itch-Related Magnetic Fields
and Itch-Related Magnetic Fields
 Relaxometry: A Comparison to Magnetoencephalography
Relaxometry: A Comparison to Magnetoencephalography
 for Measuring MEG Signals
for Measuring MEG Signals




Related posts:
Decoding and Brain Machine Interfaces Based on Electromagnetic Oscillatory Activities: A Challenge for MEG
Neuroscience: A New Frontier for Magnetoencephalography?
Magnetoencephalography (fMEG)
and Itch-Related Magnetic Fields
Relaxometry: A Comparison to Magnetoencephalography
for Measuring MEG Signals
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
