Fig. 1.1
Course of the pudendal nerve and its terminal branches
Superficial branches supply the skin of the perineum and posterior scrotum. The rectal nerves innervate the external anal sphincter. The dorsal nerve of the penis penetrates the suspensory ligament of the penis and supplies the penile shaft skin and glans penis and striated urethral sphincter [6, 7].
The deep branches of the perineal nerves give motor supply to the striated urethral sphincter, ischiocavernosal muscles and bulbospongiosus muscle [8, 9]. Contraction of the ischiocavernosus muscles produces the rigid–erection phase. Rhythmic contraction of the bulbocavernosus muscle is necessary for ejaculation.
Takenaka found that the distance from the lowest point of the endopelvic fascia to the point where the sphincteric branch of the pudendal nerve entered the urethral rhabosphincter was a mean of 5.5 mm (range 3–8 mm) [10]. Narayan similarly found branches from the dorsal penile nerve innervating the rhabdosphincter were in close proximity (3–13 mm) to the prostatic apex. Thus these branches can potentially be injured during the apical dissection of the prostate [7]. Direct injury to the branch of the pudendal nerve as it extends into the dorsal penile nerve has been reported with the perineal approach [11] (Fig. 1.2).
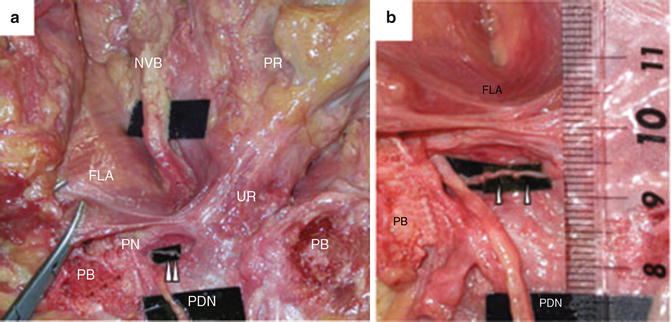
Fig. 1.2
Relationship between the fascia levator ani (FLA) and the sphincteric branch from the pudendal nerve (in the right pelvis). (a) Dissection around the membranous urethra in the fresh cadaver. A small branch from the pudendal nerve entered the rhabdosphincter, representing the sphincteric branch; (b) Close-up photograph of the sphincter branch in panel (a). The distance from the lowest point of the fascia levator ani (FLA) to the entry point into rhabdosphincter was 7 mm. PDN penile dorsal nerve, PN pudendal nerve, PB pubic bone, UR urethra, PR prostate, NVB neurovascular bundle; arrowhead, sphincteric branch from the pudendal nerve. Reprinted from Takenaka A et al. A novel technique for approaching the endopelvic fascia in retropubic radical prostatectomy, based on an anatomical study of fixed and fresh cadavers. BJUI (2005); 95(6):766–71, with permission from John Wiley and Sons
It is generally accepted that the pudendal nerve has purely somatic function and that the cavernous nerve only regulates autonomic function. Consistent with their functions, the pudendal nerve is located inferior to the pelvic diaphragm in the ischio-anal fossa and the pelvic plexus lies superior to the pelvic diaphragm. Cadaveric studies and more recently computer-assisted dissections of foetal cadavers have shown that the pelvic structures responsible for sphincter continence and sexual function receive dual innervation from the autonomic supralevator (cavernous nerve) and the somatic infralevator pathways (dorsal penile nerve) with communicating branches between them [3, 12–14]. This suggests possible plasticity in the supply of nerves for penile erection.
The Pelvic Plexus and Neurovascular Bundle (NVB)
Sympathetic fibres of the superior hypogastric plexus, the sacral sympathetic chain ganglia and parasympathetic fibres of the pelvic splanchnic nerves, as well as somatic afferents, feed into the pelvic plexus. This is the main coordinating centre for pelvic autonomic innervation.
In 2004, our team at the Royal Melbourne Hospital performed a series of detailed anatomical studies on 12 adult cadavers [15]. Four of the 12 cadaver specimens were hemisected. The fascial sheaths overlying the sacral nerve trunks were excised. The parasympathetic pelvic splanchnic nerves were identified branching from the anterior sacral nerve trunks S2–S4 and dissected as they descended anteriorly to pass beneath the lateral border of the rectum. The rectum was transected at the level of S1–S2 and its anterior surface reflected from the posterior aspect of the prostate. Extreme care was taken when freeing the anterior surface of the rectum from the prostate, as dispersed throughout the separating fascial layers (collectively known as Denonvilliers’ fascia) and adipose tissue laid the neurovascular tissue under investigation. Both layers of Denonvilliers’ fascia (prostatic and rectal) were left on the posterior surface of prostate to ensure an adequate safety margin for preserving neurovascular tissue. The mobilized rectum was displaced posteriorly, revealing the underlying pelvic splanchnic nerves entering the pelvic plexus.
In the remaining eight cadaver specimens, the dissection sequence was altered with an en bloc pelvic resection adopted. Pelvic blocks were excised from the remaining eight cadaveric specimens. Both lower limbs were amputated at the upper extremity of the thigh and the abdomen transected 1 cm above the iliac crest (with previous removal of lower abdominal viscera). The lateral and posterior aspects were sectioned exposing the levator ani musculature and the posterior wall of the rectum. Remnants of the bony pelvis and perirectal adipose tissue were removed, clearly exposing the lateral surface of the levator ani musculature and the posterior and lateral walls of the rectum, respectively. The rectum was reflected and displaced posteriorly, exposing Denonvilliers’ fascia with an investing layer of perirectal fat. When reflecting the rectum, care was taken to preserve its neurovascular supply. The perirectal fat was excised and Denonvilliers’ fascia carefully dissected, with the anatomy of Denonvilliers’ fascia and its relationship to the interposed and underlying neurovascular tissue noted. Loose connective tissue and adipose tissue surrounding the prostate, seminal vesicles and bladder was removed, rendering them clearly visible. This dissection sequence enables the pelvic plexus to be dissected in full, and exposed the additional branches innervating the bladder, seminal vesicles, anterolateral prostate and levator ani musculature. Although innervation to these structures was noted, the focus was directed on documenting the nerve branches descending within the NVB. The levator ani musculature was excised to varying extents, and the lateral pelvic fascia resected to gain greater exposure of the NVB and anterolateral surface of the prostate. The prostatic apex, external urethral sphincter, perineal membrane and membranous urethra were exposed. The cavernosal nerves were traced to the corpora cavernosa.
With both methods of dissection, the pelvic plexus and NVB were meticulously dissected under magnification (×6) with the interposed adipose tissue and fascial layers carefully removed. The relationship of the NVB to surrounding structures and the target organs of its constituents was documented.
The fenestrated pelvic plexus is a retroperitoneal, rhomboid shaped collection of ganglia with a longitudinal diameter of 3–5.5 cm. It is situated on the lateral surface of the rectum—separated by the pararectal fascia and 1–2 cm of perirectal adipose tissue. It extends as far as 1.5 cm posterior to the dorsal edge of the rectum and 1 cm superior to the rectovesical pouch (Fig. 1.3).
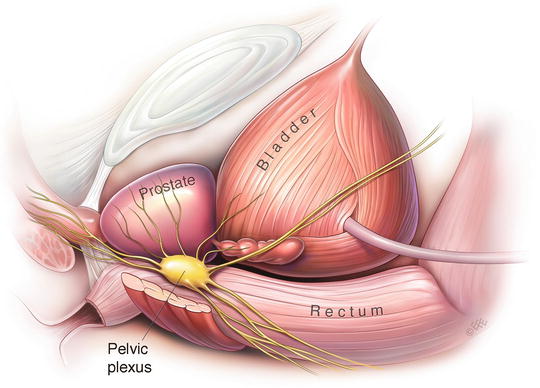
Fig. 1.3
Illustration of the left pelvic plexus and its relations
We found that there is a quantitative relationship between the size and mass of neural tissue within the pelvic plexus and the number of nerve branches within its projections. The branches of the pelvic plexus form three major projections: (1) anterior, extending across the lateral surface of the seminal vesicle and the inferolateral surface of the bladder; (2) antero-inferior, extending to the prostatovesical junction and obliquely along the lateral surface of the prostate; and (3) inferior, running between the rectum and the posterolateral surface of the prostate, forming the neural constituents of the NVB (Fig. 1.4).
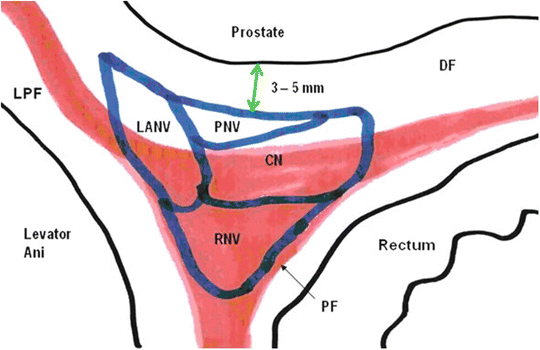
Fig. 1.4
The fascial compartments of the NVB. RNV neurovascular supply to the rectum, DF denonvilliers’ fascia, PF pararectal fascia, LPF lateral pelvic fascia, LANV neurovascular supply to levator ani, PNV neurovascular supply to the prostate, CN cavernosal nerves. Adapted from images courtesy of Department of Anatomy, The University of Melbourne
The pelvic plexus is closely associated with branches of the inferior vesical vein and artery. These large vessels are predominantly in a sagittal plane that is superimposed on the lateral surface of the pelvic plexus. On removing investing adipose and connective tissues, these vascular and neural (pelvic plexus) structures generally lay in distinct separable layers posteriorly, only to converge at the level of the pelvic plexus projections.
Course of the NVB (Fig. 1.5)
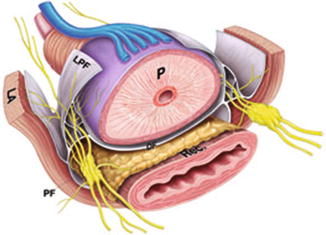
Fig. 1.5
Illustrative representation of periprostatic autonomic innervations. Denonvillers’ Fascia (DF), Levator ani (LA), lateral prostatic fascia (LPF), pararectal fat (PF), rectum and prostate (P), rectum (Rec). Reprinted from Costello et al. Immunohistochemical study of the cavernous nerves in the periprostatic region. BJUI 2010;107:1210–15, with permission from John Wiley and Sons
The inferior projection of the pelvic plexus unites with several vessels to form a prominent NVB, where it descends between the rectum and posterolateral border of the prostate.
In all 24 dissections, the plexus of nerves running within the NVB branch from the postero-inferior aspect of the pelvic plexus are 0.5–2 cm inferior to the level of the tip of the seminal vesicle.
The number of macroscopic nerves present varies, with 6–16 noted. On branching from the pelvic plexus these nerves are spread significantly, with up to 3 cm separating the anterior- and posterior-most nerves.
The nerves located most anteriorly are intimately associated with the seminal vesicle, coursing along the posterolateral surface, while the nerves located posteriorly run dorsal to the posterolateral verge of the seminal vesicle. Generally, most of the NVB descends posteriorly to the seminal vesicle. The nerves converge en route to the mid-prostatic level, forming a more condensed NVB, only to diverge once again when approaching the prostatic apex.
The nerves of the NVB are intimately associated with vessels branching from the inferior vesical vein and artery. As these vessels course distally toward the prostatic apex numerous terminal branches are given off which, in most cases, mimic the course of the nerves.
The nerves running in the NVB innervate the corpora cavernosa, rectum, prostate and levator ani musculature. Nerves running in the NVB pass through slit-like openings in the lateral pelvic fascia to innervate the superior and middle sections of the levator ani musculature. Many nerve and vascular branches pierce the lateral pelvic fascia distally to supply the inferior portion. The nerves innervating the posterior aspect of the prostate are intimately associated with capsular arteries and veins of the prostate. These structures penetrate the prostatic capsule along its base, mid-portion and apex.
The cavernosal nerves and several small vessels pierce the urogenital diaphragm posterolateral to the prostatic apex. At this level the clearly visible cavernosal nerves divide into numerous small branches that descend along the posterolateral aspect of the membranous urethra, before penetrating the posterior aspect of corpora cavernosa.
These findings confirmed the course of the cavernosal nerves in the posterolateral groove between the prostate and rectum as described in the seminal work on foetal cadaveric dissections of Walsh and Donker [16].
Functional Compartmentalization of the NVB
The constituents of the NVB are organized into three functional compartments (Fig. 1.4). The neurovascular supply to the rectum is generally in the posterior and posterolateral sections of the NVB, running within the leaves of Denonvilliers’ and pararectal fasciae. The levator ani neurovascular supply is in the lateral section of the NVB, descending along and within the lateral pelvic fascia. The cavernosal nerves and the prostatic neurovascular supply descend along the posterolateral surface of the prostate, with the prostatic neurovascular supply most anterior. Part of this anterior compartment runs ventral to Denonvilliers’ fascia. The functional organization of the NVB is not absolute and is less pronounced proximally at the levels of the seminal vesicles and the prostatic base. In addition to the nerves descending within the NVB, a scattering of nerves extends from the medial margin of the NVB to the prostatic midline. The deepest nerves (from an anterior aspect) innervate the anterior surface of the rectum at the level of the prostatic apex. The more superficial nerves descend posterior to the prostatic apex and merge laterally with the NVB.
This significant discovery of distinct fascial compartments within the NVB showed that the synonymous use of the terms NVB and cavernosal nerves in previous publications was not appropriate.
Sural nerve grafts to treat post radical prostatectomy erectile dysfunction were in vogue at the time of this study. The finding of additional innervation beyond that of the carvenosus nerves within the NVB meant that one could no longer just anastomose the sural nerve graft to any intraoperatively identified nerve fibre in the NVB.
Are There Parasympathetic Nerves Outside of the NVB?
The concept of the well-defined NVB posterolateral to the prostate was challenged subsequently by several investigators. Kiyoshima found a recognizable NVB in the posterolateral region of the prostate in only 48 % of radical prostatectomy specimens, while the rest of the specimens showed vessels and nerve trunks sparsely spread from the lateral aspect of the prostate to the anterior, without a definite bundle formation [17]. Eichenburg found in his study of radical prostatectomy specimens that a quarter to one-fifth of nerves are found on the ventral surface of the prostate [18]. In his study of foetal and adult cadavers, Lunacek found that the growth of the prostate influences the topographic anatomy of the cavernous nerves, with the nerves displaced more anteriorly and dispersed along the convex surface of the prostatic capsule [19]. These findings supported the cadaveric studies by Takenaka, who found that parasympathetic nerve components joined the NVB in a spray-like distribution at multiple levels [20].
These authors speculate that these anteriorly placed nerves contribute to the pro-erectile cavernous nerves. However, no study ever showed the actual function of these nerves. Despite this lack of evidence, some institutions developed nerve-sparing techniques aimed at preserving these structures. This new nerve-sparing approach, dubbed the “Veil of Aphrodite” technique by Menon’s group in Detroit or curtain dissection by Lunacek, releases the lateral prostatic fascia high on the anterolateral margins of the prostate above the midpoint of the prostate [19, 21].
Immunohistochemical Study of the Cavernous Nerves in the Periprostatic Region
Our team felt there was little anatomical evidence to justify this approach, given that based on our previous work, the higher placed nerves were most likely destined to innervate the prostatic stroma and not the cavernosal tissue of the penis.
To provide more evidence for the different nerve-sparing approaches, we embarked on our study focused on characterizing the position and nature of the autonomic nerves surrounding the prostate using specific immunohistochemical stains to clarify the likely functionality of the more anteriorly located nerves [22].
Four blocks of pelvic tissue from the hemisectioned pelves of two embalmed and two fresh cadavers were serially histologically sectioned, from the prostatic base proximally to the apex distally. The hemisected pelvic blocks were then divided into 4-mm sections and embedded into paraffin for histological analysis. All of these initial slides were stained with haematoxylin and eosin (H&E) and, based on a primary analysis, 50 representative blocks from only the fresh cadavers were selected. From these 50 blocks, a further two slides were cut at 3 μm and mounted from each block to undergo immunohistochemical staining using both tyrosine hydroxylase (sympathetics) and neuronal nitric oxide synthase (nNOS) (parasympathetics).
To localize the parasympathetics an antibody directed against nNOS was used. nNOS is a 150-k Da protein that is found in peripheral parasympathetic nerves and catalyzes the formation of nitric oxide. Nitric oxide is released by parasympathetics and is a potent vasodilator, implicated in the physiology of the male erection.
To localize the sympathetic nerves, we used a primary antibody directed against tyrosine hydroxylase. Tyrosine hydroxylase is the rate-limiting enzyme in the synthetic pathway of norepinephrine, which is a neurotransmitter found in peripheral sympathetic nerves and their associated ganglia.
Each slide was analyzed according to nerve fibre number, type (somatic, parasympathetic or sympathetic) and position relative to the prostate. The analysis allocated nerves as they were observed to a particular sector as shown (Fig. 1.6).
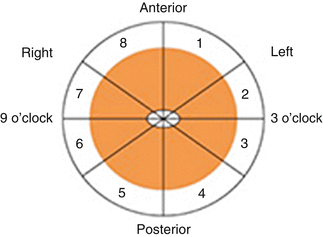
Fig. 1.6
Method of sector analysis. Reprinted from Costello et al. Immunohistochemical study of the cavernous nerves in the periprostatic region. BJUI 2010;107:1210–15, with permission from John Wiley and Sons
Results
Parasympathetic nerves accounted for 43.3 % of all nerve fibres present at the prostatic base, increasing to 45.5 % at the apex. Parasympathetic nerves found ventrally above the 3–9 o’clock level accounted for only 4 %, 5 % and 6.8 % of the total number of nerves at the base, mid-prostate and apical regions, respectively.
The proportion of sympathetic nerves was relatively constant from base to apex: 38.7–39.1 %. Sympathetic nerves found above the 3–9 o’clock level represented ≈15 % of the total number of nerves at any given prostatic level.
Somatic nerves corresponded to 18 %, 16.5 % and 15.5 % of the total nerve fibres at the base, mid-prostate and apex, respectively. Somatic nerves found above 3–9 o’clock level represented 7.5 % of the total nerves at each level of the prostate.
About a quarter (27.4 % base, 26.5 % mid-prostate and 29.6 % apical) of all the nerve fibres identified were found on the anterior half of the prostate, above the 3–9 o’clock level.
Parasympathetic Nerve Fibre Distribution (Table 1.1)
Table 1.1
Distribution of parasympathetic nerve fibres as a percentage of total parasympathetic nerves fibres at that level
Position | In NVBa | Above 3–9 o’clockb |
|---|---|---|
Prostate base | 69.4 | 9 |
Mid-prostate | 65.3 | 11.1 |
Prostate apex | 57 | 15 |
At the prostatic base, parasympathetic nerves accounted for only 14.3 % of the nerves located on the anterior aspect of the prostate. However, these nerves correspond to only 9 % of the total number of parasympathetic nerves found at the base. A significant proportion of the parasympathetic nerves at the base were localized to the region of the NVB (sectors 4 and 5). At this level, 69.4 % of the parasympathetic nerves were found within the anatomically defined NVB.
Similarly, at the mid-prostate, 65.3 % of the parasympathetic nerves were found inside the NVB. Only 11.1 % of parasympathetics at this level were found above the 3–9 o’clock junction, which, as a proportion of the total nerves at this level, represents 18.8 % of those nerves found on the anterior aspects of the prostate.
At the apex, 57 % of all parasympathetic fibres were found within the region of the NVB, with 15 % found above the 3–9 o’clock level. Importantly, the proportion of parasympathetic nerves found within the NVB dropped slightly from the base to the apex.
The fascial architecture of the posterolaterally located NVB was also analyzed and in 18 of the 32 slides examined the NVB exhibited a fascial architecture with three separate compartments as previously described. Figure 1.7 illustrates the compartmental architecture found within the NVB on a slide. Three compartments containing nerves and blood vessels can be clearly seen, the most medial of which conveyed a significant proportion of the parasympathetic nerve fibres.
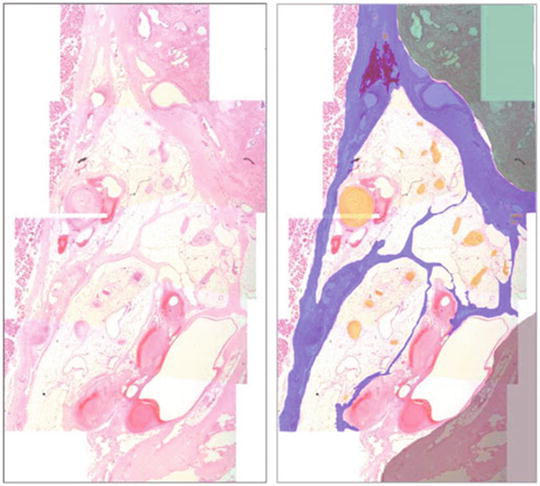
Fig. 1.7
Compartmental architecture of the neurovascular bundle (mid-prostate). Left: haematoxylin and eosin slide showing compartmental neurovascular bundle architecture. Right (overlay): prostate (green), fascial bands (blue), nerves (yellow), pararectal tissue (grey), levator ani musculature (spotted pink). Reprinted from Costello et al. Immunohistochemical study of the cavernous nerves in the periprostatic region. BJUI 2010;107:1210–15, with permission from John Wiley and Sons
Do the Anterior Parasympathetic Nerves Innervate the Corpora Cavernosa?
Our immunohistochemical staining study suggests that the anteriorly placed parasympathetic nerve fibres are likely destined for innervation of the prostatic stroma rather than the penile corpora cavernosum. This observation is supported by two pieces of evidence.
First, the total number of visible nerve fibre was smaller at the prostatic apex as compared to the base. This decrease in the number of fibre has been reported by some authors and may be related to a significant proportion of nerves penetrating the prostate to innervate the gland itself [17, 18, 23]. In the fresh cadavers in this study, 134 nerve fibres were located at the base as opposed to 115 at the apex. Of these, the absolute number of parasympathetic nerves found on the anterior half of the prostate at the apex was only eight—further supporting this hypothesis of nerves entering the prostate as they course along it.
Second, the fascial architecture of the NVB itself supports the view that prostatic innervation is the role of these anterior nerve fibres.
This study has confirmed the compartmental structure of the NVB shown in our earlier work with 12 micro-dissected cadavers [15].
Apically the absolute number of the parasympathetic fibre above the 3–9 o’clock junction increased slightly. This is consistent with studies that show the cavernous nerves ascending to assume a higher position distal to the apex. Takenaka et al. claimed the cavernous nerves assume a higher 2–10 o’clock position at the apex [24]. This study did not confirm his findings. However, careful dissection and ligation of the dorsal venous complex is recommended to preserve the neural anatomy.
Nerve Quantification: Numerical vs. Surface Area
Cross-sectional area may be a more appropriate measure of the periprostatic nerve distribution since nerve bundle quantification does not account for bundle size. To further elucidate the topography and function of the periprostatic nerves, we investigated the precise periprostatic nerve distribution in cadaveric specimens by calculating the cross-sectional area of nerve bundles [25].
Hemipelves from five fresh and two embalmed male cadavers were serially sectioned, and 4 μm serial sections representing the prostate base, mid and apical regions were prepared into slides. Mounted slides were stained with haematoxylin and eosin and digitized with a high resolution Mirax Scanner (Zeiss®) with a 20× objective. Digital sections were covered with an overlay dividing the hemiprostates into six equal sectors at a 30° angle and numbered 1–6 clockwise and counterclockwise (Fig. 1.8). Sector 5 corresponded to the posterolateral region, where Walsh identified the prominent NVB. The 13 hemiprostates were treated as independent units.
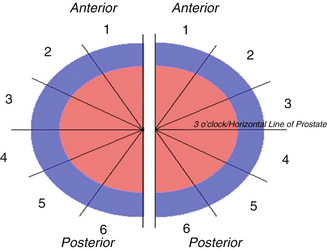
Fig. 1.8
Sector analysis used to describe nerve bundle distribution around prostate. Each sector is 30° apart from central point. From Clarebrough EE, Challacombe BJ, Briggs C, Namdarian B, Weston R, Murphy DG, Costello AJ. Cadaveric analysis of periprostatic nerve distribution: an anatomical basis for high anterior release during radical prostatectomy? J Urol. 2011 Apr;185(4):1519–25. Reprinted with permission from Elsevier
Upon analysis the periprostatic nerve bundles between the lateral prostatic fascia and Denonvilliers’ fascia posterior, and the prostatic capsule were identified and counted for all six tissue sectors. All sections were viewed using Mirax Viewer (Zeiss) software at ×5 magnification. Nerve bundles were classified as periprostatic (between the lateral pelvic fascia and capsule) or capsular (within the fibromuscular capsule). When identified, nerve cell bodies noted within sections were considered the same as nerve bundles. Cross-sectional area was calculated for periprostatic and capsular nerve bundles in each sector.
In all specimens analyzed a total of 1,199, 1,065 and 1,017 periprostatic nerve bundles were counted at the prostate base, mid part and apical regions, respectively. Total neural cross-sectional area in the base, mid part and apical periprostatic regions was 24.7, 19.7 and 13.7 mm2, respectively.
Significantly more nerve bundles were counted in the periprostatic region than in the capsule of all regions. A total of 465, 478 and 426 capsular nerve bundles were recorded in the base, mid and apical regions with a periprostatic-to-capsular ratio of 4.1, 2.4 and 3.4, respectively. The cross-sectional area of all periprostatic nerve bundles was calculated for each of the 6 sectors to determine the topographical distribution of neural tissue with respect to the prostate. The maximum neural cross-sectional area of any single sector across all specimens was 19.1 mm2 in sector 5 in the base of the prostate and the minimum was 0.2 mm2 at the sector 1 base and the sector 6 mid zone.
Sector 5, the classic location of the NVB, had a significantly higher proportion of neural cross-sectional area than all other sectors at the base, mid and apical prostate. Sectors 1 and 2 had low total neural cross-sectional areas but this increased from the base toward the apex (Table 1.2).
Table 1.2
Periprostatic nerve bundle and neural cross-sectional area per sector
Cross-sectional area | |||||||
|---|---|---|---|---|---|---|---|
– | Section 1 | Section 2 | Section 3 | Section 4 | Section 5 | Section 6 | Totals |
Periprostatic nerve bundles | |||||||
Base (mm2) | – | ||||||
Total | 0.2 | 0.4 | 0.8 | 3.2 | 19.1 | 1.0 | – |
Median (range) | 0.0 (0.0–0.1) | 0.0 (0.0–0.1) | 0.0 (0.0–0.2) | 0.1 (0.0–1.2) | 0.8 (0.4–4.5) | 0.0 (0.0–0.8) | 24.7 |
Midzone (mm2) | – | ||||||
Total | 0.4 | 0.5 | 0.7 | 1.1 | 16.7 | 0.2 | – |
Median (range) | 0.0 (0.0–0.1) | 0.0 (0.00–0.02) | 0.0 (0.0–0.2) | 0.1(0.0–0.4) | 0.8 (0.4–4.8) | 0.0 (0.0–0.1) | 19.7 |
Apex (mm2) | – | ||||||
Total | 0.4 | 1.0 | 0.8 | 1.7 | 9.5 | 0.3 | – |
Median (range) | 0.0 (0.0–0.1) | 0.0 (0.0–0.2) | 0.1 (0.0–0.2) | 0.1 (0.0–0.7) | 0.7 (0.3–1.5) | 0.0 (0.0–0.1) | 13.7 |
p value | 0.35 | 0.14 | 0.84 | 0.19 | 0.21 | 0.45 | – |
Neural tissue | |||||||
Median % (range) | – | ||||||
Base | 0.2 (0.0–4.7) | 1.2 (0.0–11.4) | 4.6 (0.9–9.6) | 10.3 (3.5–26.7) | 77.4 (62.4–94.2) | 1.0 (0.0–12.3) | – |
Mid part | 1.9 (0.0–10.1) | 2.7 (0.0–8.6) | 3.0 (0.0–19.4) | 5.3 (0.0–18.8) | 82.2 (63.7–99.3) | 0.1 (0.0–5.5) | – |
Apex | 1.8 (0.0–13.3) | 4.8 (0.2–27.2) | 4.6 (0.0–28.8) | 6.7 (0.0–43.6) | 75.5 (37.8–89.7) | 0.0 (0.0–8.6) | – |
The highest median percent of neural tissue cross-sectional area was in sector 5 with 77.4 %, 82.2 % and 75.5 % of total periprostatic neural cross-sectional area at the prostate base, mid gland and apex, respectively (Fig. 1.9).
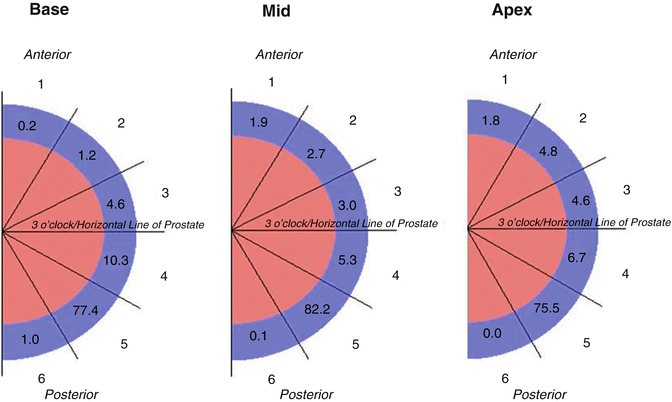
Fig. 1.9
Values in blue areas indicate median percent of periprostatic neural cross-sectional area per sector at prostate base, mid part and apex. From Clarebrough EE, Challacombe BJ, Briggs C, Namdarian B, Weston R, Murphy DG, Costello AJ. Cadaveric analysis of periprostatic nerve distribution: an anatomical basis for high anterior release during radical prostatectomy? J Urol. 2011 Apr;185(4):1519–25. Reprinted with permission from Elsevier
The percent of neural tissue surrounding the anterior part of the prostate was far lower than that around the posterior part. However, this varied and increased from base through to apex with 6.0 % at the base, 7.6 % at the mid part and 11.2 % at the apical anterior prostate.
Ganzer et al. similarly investigated the distribution of nerve bundles of prostatectomy specimens using the cross-sectional area of nerve bundles or planimetry rather than simple nerve quantification [26]. They reported that only 7.2 %, 19.4 % and 14.2 % of neural tissue surrounded the anterior part of the prostate at the base, mid zone and apex, respectively. Also, there was a significant decrease in total nerve surface area from the base over the middle toward the apex of the prostate as the periprostatic nerves penetrated the capsule to directly innervate the prostate.
The limitations of these studies were that, while the distribution of the neural tissue could be quantified, the contribution to erectile physiology was unknown because of the use of non-specific staining. Ganzer et al. addressed this problem with their recent detailed characterization of the topographic distribution of periprostatic nerves, including immunohistochemical differentiation of pro-erectile parasympathetic from sympathetic nerves of 49 radical prostatectomy specimens [27].
They confirmed our previous findings that more than 75 % of neural tissue is distributed in the dorsolateral position, with less than 15 % of parasympathetic neural tissue found ventrally. In addition, they found that the proportion of parasympathetic tissue ventrally did not increase towards the base. They concluded that high incision might help preserve more parasympathetic nerves at the base and the middle but is of little help at the apex.
The drawback of all these anatomical studies is that none are able to confirm if these anteriorly placed parasympathetic nerves contribute to erectile function. Kaiho et al. measured the increase in intracavernosal pressure after intraoperative electrical neurostimulation of the lateral periprostatic nerves by a small bipolar electrode [28]. This small study demonstrated that every electrostimulation of the nerves found between the 1 and 5 o’clock positions on the lateral surface of the prostate resulted in an increase in cavernosal pressure. Of note the mean pressure response was most powerful for 5 o’clock stimulation and decreased with stimulated points further from the 5 o’clock position. While this is suggestive that the anterior periprostatic nerves contribute to erectile function, artificial electrical stimulation may be too crude a technique to provide anatomical proof of function.
As long as the physiology of these nerves is not clearly verified by functional studies, one should undertake every effort in nerve-sparing prostatectomy procedures to preserve as many of these nerves as possible. With this in mind, we suggest that a slightly modified incision technique be used to dissect the NVBs off the prostate, making a sweeping incision that climbs anterior up to the 1 o’clock position at the apex but not in the more proximal regions of the gland.
Relationship of Pelvic Fascial Layers to Nerve-Sparing Prostatectomy
Having described the location of the CN in relation to the NVB and periprostatic area, understanding the location and distribution of the fascias around the prostate and their relation to the NVB becomes key to achieve the desired degree of nerve sparing during the radical prostatectomy (RP). There are four main fascial layers surrounding the prostate and NVB, which serve as surgical landmarks that will help determine the layers that should be incised in order to remove or preserve different amounts of cavernous nerve tissue during RP.
Endopelvic Fascia (EPF)
The endopelvic fascia has two components: parietal and visceral. The parietal EPF covers the levator ani lateral to the fascial tendinous arch of the pelvis. The visceral EPF covers the detrusor apron and anterior surface of the underlying prostate. The parietal and visceral EPF are fused at the lateral side of the prostate and bladder. This fusion site is sometimes recognized as a whitish line and is called the “arcus tendineus fascia pelvis” (ATFP) [29]. The ATFP runs from the puboprostatic ligament to the ischial spine.
Conventional wisdom is that the EPF should be incised lateral to the ATFP to avoid unnecessary haemorrhage from prostatovesical veins [30]. Takenaka and colleagues showed in cadaveric studies that preserving the fascia of the levator ani helps to protect the levator ani muscle, rhabdosphincter and pudendal nerve branches to the rhabdosphincter [10]. This concurs with what other pioneering prostatectomy surgeons advocate—that the correct location to incise the fascia is medial to the fascial tendinous arch—thus leaving the levator ani muscle covered by its fascia [31].
Occasionally, part of the endopelvic fascia over the prostate coalesces with that lying over the levator ani fascia at its deepest point of reflection. This fusion of layers needs to be detached for an adequate incision of the endopelvic fascia. Applying countertraction on the prostate facilitates identification and dissection of the contour of fascial reflection where the endopelvic fascia forms a cul-de-sac between the levator ani fascia laterally and the prostatic fascia medially. It is easier to identify and correctly incise this cul-de-sac when this dissection is initiated close to the base of the prostate (as opposed to close to the apex) where the endopelvic fascia seems to be thinner and the space between levator ani and the prostatic fascia is wider.
Periprostatic Fascia
The fascia on the outer surface of the prostate has been given several different names by different authors. It has been referred to as periprostatic fascia by Myers and Villers [32], Stolzenburg et al. [33] and Tewari et al. [34]; as lateral pelvic fascia by Costello et al. [15], Takenaka et al. [10] and Walsh and Partin [35]; as parapelvic fascia by Graefen et al. [36] and Budaus et al. [37]; and Menon et al. [38] and Secin et al. [39] have used the term prostatic fascia.
This fascia is not a discrete single-layered structure stretching over the lateral surface of the prostate. Often it is multilayered structure over the prostate, consisting of both collagenous and adipose tissue elements. Walz has done an excellent review on the pelvic fascias and their relationship to the NVB [40]. Similarly, for ease of review, we will use the term periprostatic fascia (PPF) to signify all fascias on the prostate that are external to the prostate capsule. The PPF covering the prostate can now be subdivided into three basic elements according to location.
Anterior Periprostatic Fascia (Visceral Endopelvic Fascia)
This represents the visceral endopelvic fascia discussed earlier in this section. It is associated with the anterior surface of the prostate from approximately the 10-o’clock to 11-o’clock positions to the 1-o’clock to 2-o’clock positions, where it covers the detrusor apron, dorsal vascular complex (DVC), and is fused in the midline with the anterior fibromuscular stroma of the prostate.
Lateral Periprostatic Fascia
Once the endopelvic fascia is opened lateral to the fascial tendinous arch of the pelvis and the levator ani muscle is deflected laterally, the outermost fascial layer on the lateral surface of the prostate is the levator ani fascia (LAF).
Under this layer lies the prostatic fascia. The PF is fused anteriorly with the puboprostatic ligaments/dorsal venous complex, coming into intimate contact with the prostate pseudocapsule more laterally [21].
Both of these layers (LAF and PF) constitute PPF for the operating surgeon. The outer LAF covers the underlying, laterally distributed levator ani muscle and therefore forms the lateral boundary of the axial triangular space occupied by the bulk of the cavernous nerves and vessels, and varying amounts of fat. It extends posteriorly over the muscle fibres and continues over the lateral aspect of the rectum to form the so-called pararectal fascia (separating the rectum from the levator ani). The medial boundary of this triangle is formed by the inner PF and underlying prostate pseudocapsule and the posterior boundary by Denonvilliers’ fascia [15].
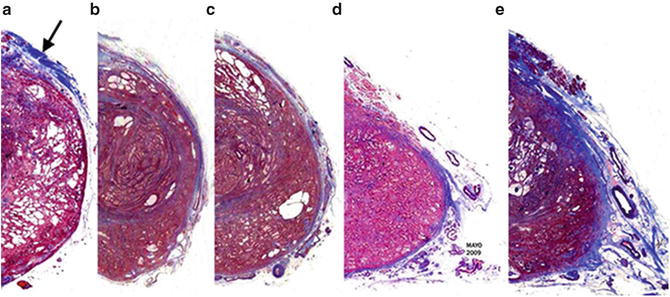
The relationship between the prostate capsule and the lateral PF may differ depending on interindividual variations (Fig. 1.10). Kiyoshima et al. found that in 52 % of all cases, the LAF, which they called “lateral pelvic fascia,” does not adhere to their prostate “capsule” [17]. In the cases just cited, the apparent space consisted of loose connective and adipose tissue referred to as areolar tissue. Furthermore, in such cases the NVB could not be identified as a distinct structure but was spread out over the lateral prostatic surface. In the remaining 48 %, LAF was fused with their prostate capsule, and no areolar tissue was seen between the layers. An exception to this fusion was only seen at the posterolateral angle of the prostate, and the NVB was identified there as a distinct bundle.
 Challenging Cases in Robotic Radical Prostatectomy
Challenging Cases in Robotic Radical Prostatectomy
 Postoperative Management and Preoperative Considerations: Urinary Incontinence and Anastomotic Stricture
Postoperative Management and Preoperative Considerations: Urinary Incontinence and Anastomotic Stricture
 Patient Positioning; Incision/Port Placement
Patient Positioning; Incision/Port Placement
 Robotic-Assisted Radical Prostatectomy: A Surgical Guide to the Extraperitoneal Approach
Robotic-Assisted Radical Prostatectomy: A Surgical Guide to the Extraperitoneal Approach
 The Transperitoneal Robotic-Assisted Radical Prostatectomy
The Transperitoneal Robotic-Assisted Radical Prostatectomy
 Open Radical Retropubic Prostatectomy
Open Radical Retropubic Prostatectomy




Related posts:
Challenging Cases in Robotic Radical Prostatectomy
Postoperative Management and Preoperative Considerations: Urinary Incontinence and Anastomotic Stricture
Patient Positioning; Incision/Port Placement
Robotic-Assisted Radical Prostatectomy: A Surgical Guide to the Extraperitoneal Approach
The Transperitoneal Robotic-Assisted Radical Prostatectomy
Open Radical Retropubic Prostatectomy
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
