(1)
Department of Paediatric Pathology, Great Ormond Street Hospital for Children, London, WC1N 3JH, UK
10.1 Overview
Embryonic development of the head and face is a complex process occurring during the first trimester, involving interactions between components from all three germ layers, between neural tube formation and associated head and facial development and between multiple branchial/pharyngeal arch components. For these reasons, numerous abnormalities may occur depending on the site, timing and severity of any defects in these processes. However, the general types and mechanisms of such anomalies can be reasonably grouped together, and this chapter aims to provide a brief overview of the major components of facial development in association with clinically significant fetal abnormalities. In addition, the chapter will briefly describe the approach to investigation of such abnormalities at autopsy and major associated features. For those interested, there are excellent and highly detailed accounts available of these processes from an embryological perspective, documenting the mechanisms of cellular interactions during these processes and with experimental evidence for the developmental origins of the various components, which are beyond the scope of this chapter [7, 12, 16, 21].
10.2 Developmental Aspects of the Head and Face
Development of the head, particularly of the facial structures, represents a cohesive series of events resulting in formation of several anatomical systems including the central nervous system, specialist sense organs, such as the ears, and the first parts of both respiratory and alimentary systems including the nose, pharynx and mouth. In addition, the area demonstrates complex developmental formation of bony structures, muscles and innervation to allow blinking, chewing, swallowing, talking and facial expression.
In order to achieve this final arrangement, embryonic development requires tightly controlled interactions between the numerous systems and developmental elements. The most important overriding concepts in this regard for overall head and face development are:
Formation of the underlying brain and its impact on development of both the cranium and face
The pivotal role of the neural crest for all aspects of development at this site
The important contributions of the branchial/pharyngeal arches to both facial and neck structure development
10.2.1 Neural Development
Formation of the brain proper will not be covered here; suffice to note that the brain structures proper, peripheral nervous system elements and ectodermal placodes are ultimately derived from the neural plate and neural folds, this tissue forming the brain structures proper, peripheral nervous system elements and ectodermal placodes. Furthermore, as the neural plate forms the neural tube and subsequent central nervous system, non-neuronal neural crest elements play an important contribution to the mesenchymal elements of various head and face structures, as indicated below.
The general brain regions are formed during early conception, even before the neural tube itself has closed. With further differential growth of these regions, a series of folds or flexures occur that divides the brain into segments, each forming distinct components of the mature central nervous system. Concurrently, neural crest cells from these regions are formed, which then contribute to specific regional elements such as trigeminal, facial, vestibuloacoustic and superior petrosal ganglia, in addition to other elements of the sympathetic and parasympathetic nervous system in this region. Similarly, the regional and ordered distribution of the cranial nerves and their nuclei are determined at this time [22].
10.2.2 Skull Development
The skull develops as the cranium proper from local paraxial mesenchyme, which forms the membranous frontal, parietal, squamous and occipital bones that surround the brain. The face and jaw bones are derived from neural crest components.
10.2.3 Overview of Facial Development
The structural complexity of the face is due to its development being related to the growth and folding of the brain, cranial coverings, components formed from the branchial arches (see later) and also the interrelationship with the endodermally derived foregut system [4]. The face essentially forms around the cranial end of the foregut, which is initially closed at the buccopharyngeal membrane, the junction between the endoderm and ectoderm, within a depression, the stomodeum. This area will become the mouth and the other facial elements are hence formed from the surrounding areas. The buccopharyngeal membrane breaks down, and with development of the pharyngeal arches, this part of the foregut forms the mouth and pharynx and its derivatives. Therefore, the majority of the face and neck, including the pharynx and larynx, are derived from a combination of neural fold-derived ectoderm and mesoderm from the pharyngeal arches which surround the developing foregut.
The majority of facial development in terms of formation of structures occurs around weeks 4–8 postconception. Further anatomical changes occur as a consequence of differential growth rather thatn development of new structures.
10.2.4 Branchial/Pharyngeal Apparatus
Given the importance and complexity of the development of the branchial/pharyngeal apparatus to the formation of the head and neck structures, this section will provide further detail. These primitive structures form gills in fish but contribute to many head and neck elements in the human [15]. Around week four of development, neural crest cells migrate into ridges on either side of the future head, which are separated by grooves or clefts, with four major arches (five in total) forming in humans. With subsequent growth these structures become fused and the outer surface of the face and neck is smooth.
The five pharyngeal arches are each composed of a covering of ectoderm (some also contain endoderm), with associated neural crest-derived mesenchymal elements. The arches therefore contribute to epithelial-derived structures, mesenchymal elements such as connective tissue and muscle and mesenchymal-derived vascular structures and nerves. Facial muscles are derived from branchial arch mesenchyme. Each arch is associated with a corresponding cranial nerve, explaining the innervation of face and neck structures. The overall control of this process in terms of order and regulation appears to be largely controlled by homeobox (Hox) gene expression patterns [4], but a large number of additional factors are now being recognized as potentially important, including BMPs and FGFs [13, 23].
The first arch structures form the lower jaw and midface, including the maxilla and mandible, bones of the middle ear (malleus and incus) and associated muscles and ligaments, and are associated with the trigeminal nerve.
The second arch forms the middle ear stapes and part of the temporal and hyoid bones and is associated with the facial nerve.
The third arch forms part of the hyoid and is associated with the glossopharyngeal nerve.
The fourth to fifth arches form the larynx and are associated with the vagus nerve.
Since the arches are separated by grooves, four well-defined pharyngeal pouches also therefore form, in which the primitive foregut/pharyngeal endoderm is in close proximity to the ectoderm at the base of the grooves.
The first pouch forms the tympanic membrane, cavity and mastoid antrum and Eustachian tube. The second pouch forms the tonsillar fossa, the third pouch forms the inferior parathyroid glands and thymus and the fourth pouch forms the superior parathyroid glands. The first groove therefore persists as the external auditory meatus, whilst all other grooves are obliterated, providing the normal smooth outline of the neck.
Failure of this complex process of endodermal, mesodermal and ectodermal cellular interaction, differential growth and migration may result in a range of associated anomalies, which can be readily predicted based on the knowledge of their normal embryological derivatives. For example, persistence of first groove elements may result in auricular anomalies and congenital auricular sinuses/remnants. Branchial sinuses on the lateral neck are a consequence of persistence of second groove elements. True branchial fistulas result from communications at the site of the base of the pharyngeal grooves, usually between the tonsillar clefts and lateral neck, and remnants of the second branchial groove or cervical sinus may result in branchial cysts.
Similarly, abnormal development of the various branchial arch structures themselves can result in a range of anomalies in relation to their developmental structures described above. Anomalies of the first branchial arch may be associated with abnormal development of the eyes, ears, mandible and palate, presumably a consequence of defective neural crest migration or interaction with surrounding tissues. Examples of such so called “first arch syndromes” include Treacher Collins syndrome and Pierre Robin syndrome, both of which have characteristic facial dysmorphic features. Failure of normal third and fourth arch development leads to DiGeorge syndrome with thymic aplasia and absent parathyroid glands.
10.2.5 Formation of the Central Facial Structures
The face itself is formed from several components, or facial primordia, which develop around the primitive stomodeum. The frontonasal processes develop from the neural fold and significant components, including the paired maxillary and mandibular processes, develop from the first pharyngeal arch, including the paired maxillary and mandibular processes. Neural fold mesenchyme forms around the olfactory placodes forming the medial and lateral nasal swellings, which fuse with the frontonasal prominence to form the nose. The nasal cavities form from underlying nasal sacs, and with degeneration of the oronasal membrane and subsequent formation of the palate, the nasal and oral cavities are separated, but connected by the choanae. Ectodermal epithelium in this region forms the olfactory epithelium. The paranasal sinuses develop only during late fetal life and into childhood, being small and primitive in the fetus.
The maxillary processes of the first arch fuse with the lateral nasal swellings in the midface, the epithelial junction of which (the nasolacrimal groove) develops into the nasolacrimal duct. The maxillary processes also grow medially, via the palatine shelves, to fuse together in the midline and also join with the intermaxillary segment of the frontonasal prominence/premaxilla (which also forms the central lip philtrum), to form the normal intact lip and palate (Figs. 10.1 and 10.2).
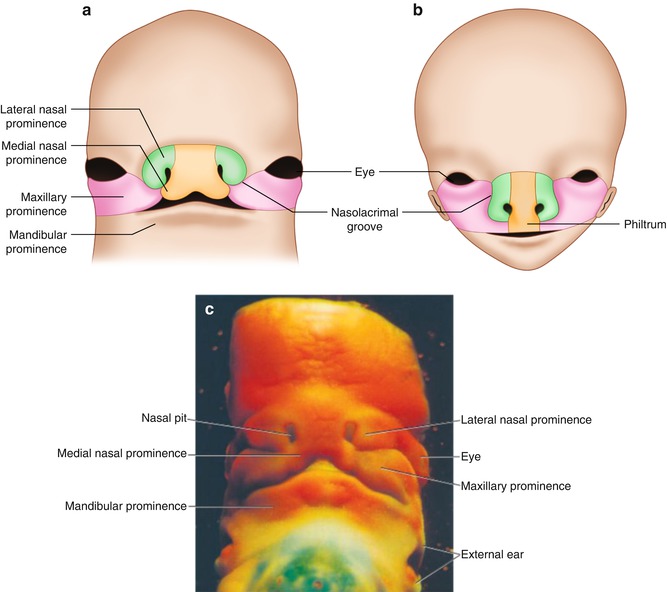
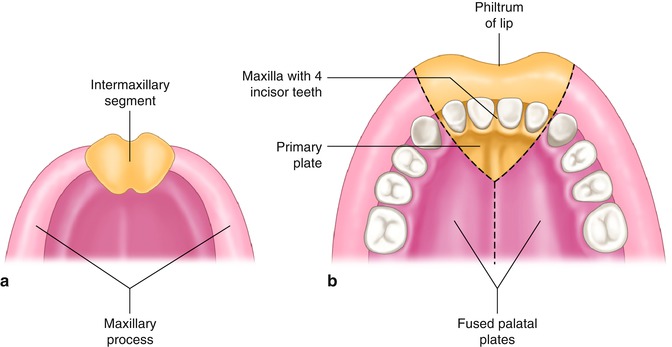
Fig. 10.1
Frontal aspect of the face. (a) Seven week embryo. Maxillary prominences have fused with the medial nasal prominences. (b) Ten week embryo. (c) Photograph of a human embryo at a stage similar to that in (a) (From Sadler [16], 12th ed, Fig 17.23 p. 277 and Fig. 17.24 p. 278)
Fig. 10.2
(a) Intermaxillary segment and maxillary processes. (b) The intermaxillary segment giving rise to the philtrum of the upper lip, the median part of the maxillary bone with its four incisor teeth, and the triangular primary palate (From Sadler [16], 12th ed, Fig 17.23 p. 277 and Fig. 17.24 p. 278)
If such elements fail to fuse normally, facial clefting of various types may result [20]. For example, if mesenchymal fusion between the maxillary process and nasal process is defective, typical anterior asymmetrical cleft lip occurs; the central philtrum area, being formed from the frontonasal process, is often intact although the nasal cavity floor is usually defective. Bilateral cleft lip (Fig. 10.3) occurs when the maxillary processes fail to fuse bilaterally with the medial nasal process. Central cleft palate is due to failure of the midline maxillary processes/palatine shelves to fuse [6]. Less commonly, more extensive fusion failure may result in severe clefts, such as an oblique cleft due to persistence of the nasomaxillary groove, and rarely, other types of facial cleft may occur due to failure of embryological development or fusion at other sites, such as midline cleft.
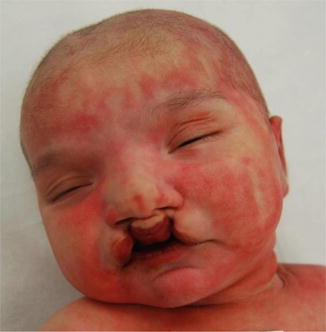
Fig. 10.3
Bilateral cleft lip with hypertelorism and a broad nasal bridge. Bilateral clefting occurs when the maxillary processes fail to fuse bilaterally with the medial nasal process
10.2.6 Tongue
The tongue forms in the floor of the primitive pharynx as the median tongue bud, with lateral tongue buds developing from the mesenchyme of the first branchial arcehes, which fuse to form the anterior part of the tongue. The posterior, pharyngeal elements of the tongue are derived from the second, third and fourth branchial arches. Therefore, sensation to the anterior two-thirds is derived from the trigeminal nerve (with taste buds supplied by the facial nerve), whereas the majority of the remainder of the tongue is supplied by the glossopharyngeal nerve. The tongue musculature, which is formed from the occipital somite myocytes, is supplied mainly by the hypoglossal nerve.
Anomalies of tongue development are usually associated with other abnormalities of branchial arch development, but in addition to lingual thyroid and lingual cysts due to thyroglossal duct remnants (see above), isolated abnormalities may occur such as abnormal fusion of the lateral tongue buds resulting in a cleft tongue.
10.2.7 Thyroid Gland
The thyroid gland forms from the pharyngeal endoderm, originating as the thyroid diverticulum which is initially connected to the tongue base by the thyroglossal duct. The duct then normally atrophies with associated thyroid descent. Persistence of elements of the thyroglossal duct may result in a pyramidal thyroid lobe inferiorly, ectopic thyroid gland, such as lingual thyroid, superiorly and thyroglossal duct remnants and cysts anywhere along the course of the tract in the midline of the neck.
10.2.8 Ears
The ears form from the otic placodes, bilateral areas of ectoderm that invaginate into the otic vesicles from which the inner ear structures of the cochlea and semicircular canals develop. The middle ear and Eustachian tube are endodermally derived from the first pharyngeal pouch, with the ossicles formed from the first and second pharyngeal arches (see above). The external auditory meatus and tympanic membrane are formed from the first pharyngeal cleft, whereas the auricle is formed from six auricular nodules (hillocks) from the first and second pharyngeal arches. In this manner, the ears are normally “low set” in the neck region in early embryonic life, but their normal position is attained with development of the other facial structures.
10.2.9 Eyes
Lateral outpouchings of the forebrain form the optic vesicles, which interact with the overlying ectoderm. In conjunction with the surrounding mesenchyme, which contributes to the choroid and sclera, these elements form the globe of the eye and anterior structures such as the lens and cornea. Recent understanding of the molecular interactions involved in eye development has demonstrated a complex communication between the various epithelial/mesenchymal components, with PAX6 being a major gene involved [2].
10.2.10 Teeth
The teeth are derived from a complex interaction between the ectoderm of the oral region and underlying neural crust mesenchyme. The epithelium initially forms dental buds that then develop into the tooth caps and subsequent outer layers by differentiation to ameloblasts, whereas the neural crest mesenchyme forms the central dental papilla and underlying odontoblasts and cementoblasts.
10.3 Approach to Autopsy Assessment of the Head and Face
Whilst defects of embryogenesis as discussed above are malformations (structural abnormalities arising as a direct result of an abnormal developmental process), facial abnormalities may also be deformations (structural abnormalities arising from localised or generalised mechanical forces acting on a forming anatomical structure) or disruptions (secondary destruction of an organ or structure that was previously normally formed). When assessing such abnormalities, it is therefore important to bear in mind the etiology of the pathology present and distinguish between a sporadic disruption and a malformation with significant recurrence risk. For example, a cleft-like disruption due to amniotic band sequence implies different future implications than orofacial clefting in an autosomal recessive syndrome with a 25 % recurrence risk.
For the majority of prenatally identified facial abnormalities, post-mortem assessment is straightforward, requiring direct visualisation and photographic confirmation only in most cases (Figs. 10.4 and 10.5). However, it should be recognized that changes that occur following death, especially with prolonged intrauterine retention, may lead to secondary alterations to facial appearances making interpretation of subtle dysmorphic features impossible and even making identification of more obvious anomalies more difficult (Figs. 10.6 and 10.7).
 Congenital Subcutaneous Mixed Venous-Lymphatic Orofacial Malformation Associated with Macroglossia: Prenatal Diagnosis with Ultrasound and Fetal MRI
Congenital Subcutaneous Mixed Venous-Lymphatic Orofacial Malformation Associated with Macroglossia: Prenatal Diagnosis with Ultrasound and Fetal MRI
 The Genetics of Facial Cleft
The Genetics of Facial Cleft
 Median Cleft Lip and Palate, Cutaneous Nasal Polyps, and Corpus Callosum Lipoma: A Case of Pai Syndrome Associated with Ventricular Septal Defects
Median Cleft Lip and Palate, Cutaneous Nasal Polyps, and Corpus Callosum Lipoma: A Case of Pai Syndrome Associated with Ventricular Septal Defects
 Acromelic Frontonasal Dysplasia (Median Cleft Face Syndrome)
Acromelic Frontonasal Dysplasia (Median Cleft Face Syndrome)
 Magnetic Resonance Imaging (MRI) in the Evaluation of the Fetal Face
Magnetic Resonance Imaging (MRI) in the Evaluation of the Fetal Face
 The Role of 2D/3D/4D Ultrasound in the Prenatal Assessment of Cleft Lip and Palate
The Role of 2D/3D/4D Ultrasound in the Prenatal Assessment of Cleft Lip and Palate




Related posts:
Congenital Subcutaneous Mixed Venous-Lymphatic Orofacial Malformation Associated with Macroglossia: Prenatal Diagnosis with Ultrasound and Fetal MRI
The Genetics of Facial Cleft
Median Cleft Lip and Palate, Cutaneous Nasal Polyps, and Corpus Callosum Lipoma: A Case of Pai Syndrome Associated with Ventricular Septal Defects
Acromelic Frontonasal Dysplasia (Median Cleft Face Syndrome)
Magnetic Resonance Imaging (MRI) in the Evaluation of the Fetal Face
The Role of 2D/3D/4D Ultrasound in the Prenatal Assessment of Cleft Lip and Palate
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
