Fig. 10.1
Pictographic essay of sonoporation for drug and gene delivery (Adapted from Servier Medical Art, www.servier.fr)
The first experiments on sonoporation date back to the 1980s, where various ultrasound (US) exposure conditions were tested blindly at frequencies ranging from the kHz to the MHz (Ferrara et al. 2007; Escoffre et al. 2013). Sonoporation has also been evaluated using high-pressure amplitude US waves. Since then and with the recent introduction of contrast agents, higher frequency US in conjunction with cavitation has been called on to induce a range of effects on cells. Extensive examinations have been carried out to evaluate the efficiency of US in combination with contrast microbubbles for inducing cellular uptake. Although these results and findings were achieved in a controlled in-vitro environment, diagnostic US scanners were also useful for therapeutic applications of sonoporation, particularly with the guidance of treatment afforded by the imaging mode.
One of the first papers that described the use of sonoporation with ultrasound and UCAs dates back to 1997 (Bao et al. 1997). Since then, several studies have been reported on this topic, but so far the exact mechanism of sonoporation responsible for permeabilization remains unrevealed. Available hypotheses suggest that acoustic phenomena, such as stable and inertial cavitation, microstreaming, microjets, and shock waves are involved in the observed permeabilization sequences. All these acoustic phenomena are induced by the oscillations or destructions of microbubbles activated by US waves. Although the results obtained from these studies demonstrate the potential of sonoporation as a therapeutic strategy, they still provide conflicting conclusions. Some studies show that inertial cavitation is required for the induction of drug uptake; whilst others show that stable cavitation is sufficient. Microbubble jetting has been observed near a cell layer, but its effect on drug uptake was not really studied. The influence of microstreaming has rarely been studied in experiments using encapsulated contrast microbubbles, but there are various theoretical approaches. Other studies show that standing waves are required for drug uptake. A few recent reports have shown that oscillating microbubbles can deform cells, which could be a trigger for drug uptake. This suggests that drug uptake can be induced by several acoustic mechanisms. It has been reported that the conclusions about the mechanisms of cell membrane permeabilization are not straightforward despite numerous papers using various cell line models, ultrasound systems and experimental environments. Current investigations report only on drug uptake, electron microscopy observations, or cell electrophysiological measurements but do not address the relationship between microbubble acoustic behavior and drug uptake.
In this chapter, firstly we will present various acoustic phenomena that originate from the interaction of ultrasound waves and microbubbles and that are likely to be involved in the permeabilization of the cell membrane. In the second part, various assumed mechanisms by which molecules or drugs extravsate through the cell membrane and enter into the cytoplasm after sonoporation will be discussed. Finally, we will give a number of examples of in-vivo applications of sonoporation.
10.2 Mechanisms of Barrier Permeabilization and Molecular Delivery
Sonoporation was first demonstrated in the late 80s where kHz frequency ultrasound was evaluated for molecular uptake. This finding was original, although the efficiency of this approach was not fully proved. Following the introduction of ultrasound contrast agents in the late 90s, microbubbles were thought to be associated with a higher molecular uptake. Indeed, microbubbles, which are known to be a pure intravascular tracer, do not cross the endothelial barrier. However, their oscillations next to the endothelial layer might induce the permeabilization of the vessel wall. At the cellular level, microbubble oscillations increase the cell membrane permeability transiently. The availability of different contrasts and various protocols led to a large diversity of results on molecular uptake and efficiency rendering the comparison between them nearly impossible. It is believed that contrast microbubbles can be used as a nuclei trigger to enhance the delivery of drugs and molecules through the biological barriers. Nevertheless, the diversity of the insonation parameters did not allow providing precise conclusions and comparisons between various experimental conditions.
Although the sonoporation efficiency in permeabilizing the cell membrane was demonstrated, mechanisms behind the action of ultrasound and microbubbles were not clearly identified. Therefore several groups were engaged in elucidating the mechanisms responsible for cell membrane permeabilization caused by ultrasound and microbubble insonation. Current investigations report only on drug uptake, electron microscopy observations, or cell electrophysiological measurements but do not address the relationship between microbubble acoustic behavior and drug uptake.
10.2.1 Acoustical Phenomena
Various microbubble acoustic phenomena have been explored and studied to determine their involvement in the sonoporation mechanisms. Microstreaming, microjets, stable and inertial cavitations have been proposed as possible candidates in the transient or permanent permeabilization of the cell membrane. All these acoustic phenomena are induced by the oscillations or destructions of microbubbles activated by US waves.
One of acoustic phenomena that might impact mechanically on the cell membrane to make it permeable is acoustic microstreaming. When a bubble is driven acoustically, it is known to generate steady vortical flows in the surrounding liquid. It has been suggested that this phenomenon, called acoustic microstreaming, plays an important role in various biological effects of ultrasound, such as hemolysis, sonothrombolysis and sonoporation. Early experimental investigations into this phenomenon go back to the work of Kolb and Nyborg (1956) and Elder (1959), where the dependence of acoustic microstreaming on certain parameters (e.g., amplitude of sound) was determined and observations of microstreaming patterns for various amplitudes and viscosities were made. Hughes and Nyborg (1962) were the first to show that bubble-induced vortex streaming can be used for disruption of cells in suspension. Pritchard et al. (1966) established that the number of breaks occurring in DNA molecules was directly related to velocity gradients of acoustic microstreaming. These effects were attributed to shear stresses exerted on cells and large molecules by the microstreaming. Further insights into this mechanism were given by experiments of Rooney (1970, 1972) on hemolysis produced by a single air bubble in an erythrocyte suspension.
The progress of experimental studies on microstreaming can be traced in works by Nyborg (1978), Liu et al. (2002), Tho et al. (2007), Wu and Nyborg (2008), Collis et al. (2010) and Wang et al. (2012). Wu et al. (2002) used a Mason horn with a 0.4 mm probe tip vibrating at 21.4 kHz. The transverse displacement amplitudes were greater than 7.8 μm and produced shear stresses on the Jurkat cell suspension due to microstreaming. The authors believe that the generated microstreaming around the probe tip was the main reason for the cell reparable sonoporation, inducing the uptake of fluorescent dextran molecules. The threshold was estimated to be 12 Pa for an exposure time of up to 7 min (Ross et al. 2002).
Shear stresses generated by relatively large bubbles have been recorded using a particle imaging velocity (PIV) system. The PIV measurements were performed for bubbles of different sizes ranging from 200 to 400 μm (Novell et al. 2011). The results showed that the flow velocity exhibited a first peak near the bubble resonance. For this bubble, the shear stress is also maximal. The experimental measurements also revealed a second peak for bubbles larger than the resonance size but with lower amplitude. Figure 10.2 demonstrates an example of PIV images showing the microstreaming generated around a bubble of 200 μm in diameter at 28 kHz and an acoustic pressure of 7 kPa.

Fig. 10.2
Acoustic microstreaming around an air bubble as recorded with PIV using ultrasound at 28 kHz and 7 kPa
First theoretical studies on bubble-induced microstreaming were performed by Nyborg (1958). In particular, his theory gives a formula for shear stresses exerted on a rigid plane by a pulsating hemispherical bubble resting on this plane. This formula was used by Rooney (1972) for estimating shear stresses experienced by cells near a pulsating bubble. To apply Nyborg’s formula to nonlinear bubble oscillations, Lewin and Bjørnø (1982) suggested combining Nyborg’s formula with the Rayleigh-Plesset equation. The combination of Nyborg’s formula with the de Jong shell model was used by Wu (2002) to estimate shear stresses generated by contrast agent microbubbles. It was pointed out by Doinikov and Bouakaz (2010c) that the use of Nyborg’s formula, which was derived for a hemispherical bubble, is not correct in the case of an encapsulated bubble. To solve this problem, they recalculated Nyborg’s formula for the case of a spherical bubble being at a distance from a rigid plane. The same result, in a simplified formulation, was obtained by Yu and Chen (2014). Numerical simulations of shear stresses produced by free and encapsulated bubbles on a rigid wall were performed by Krasovitski and Kimmel (2004) using the boundary integral method. They came to the conclusion that the bubble-induced stress is several orders of magnitude greater than the physiological stress induced on the vessel wall by the flowing blood.
There are a number of theoretical models for the velocity and stress fields of acoustic microstreaming that is developed around a bubble in an unbounded liquid. Davidson and Riley (1971) derived equations for microstreaming generated by a bubble undergoing translational harmonic oscillations. Wu and Du (1997) and Longuet-Higgins (1998) considered the case of a bubble undergoing translational and radial oscillations. Maksimov (2007) extended the result of Longuet-Higgins (1998) so as to include the effect of parametrically excited surface modes of high order. Liu and Wu (2009) applied the approach of Wu and Du (1997) to calculate microstreaming produced by an encapsulated bubble. Doinikov and Bouakaz (2010a, b) extended the results of Wu and Du (1997) and Liu and Wu (2009) by taking into account shape oscillations and removing restrictions such as the assumption that the thickness of the viscous boundary layer surrounding the bubble is much smaller than the radius of the bubble. Doinikov and Bouakaz (2010b) also derived equations for acoustic microstreaming generated by a bubble in the presence of a distant rigid wall. They showed that the presence of the wall can change the amplitude and the phase of the bubble oscillations in such a way that the intensity of acoustic microstreaming is increased considerably as compared to that generated by the same bubble in an infinite liquid.
The shear mechanism has also been investigated optically. The interaction of an oscillating microbubble with cells has been observed with a high-speed camera (van Wamel et al. 2004). The observations with the use of endothelial cells revealed a strong mechanical action of SonoVue microbubbles insonified with a single ultrasound burst at 1 MHz and 0.9 MPa acoustic pressure. This action induces oscillatory stresses that pull and push the cell membrane, termed here cellular massage, and likely trigger various cellular and intracellular responses.
Besides microstreaming and shear stresses, other acoustic phenomena related to microbubble oscillations have been regarded as possibly responsible for or involved in the sonoporation mechanisms and effective in permeabilizing the cell membrane at megahertz frequencies and at acoustic pressures as low as a few MPa. Shock waves and liquid microjets generated in the course of inertial cavitation are among such phenomena.
Although the dominant hypothesis is now that sonoporation is caused by shear stresses exerted on the cell membrane by acoustic microstreaming, there are suggestions that pressure shock waves and liquid microjets generated by cavitating bubbles make a contribution as well. Interest in the above phenomena goes back to applied hydrodynamic problems such as cavitation damage of ship propellers and hydraulic turbine blades and ultrasonic cleaning of contaminated surfaces. The literature on the subject is very expensive. There are a number of reviews, written in different years, that describe at length available experimental observations: Plesset and Prosperetti (1977), Mørch (1979), Blake and Gibson (1987), Leighton (1994), Brennen (1995) and Lauterborn and Kurz (2010). Experimental estimates show that the spherical shock wave produced by a collapsing bubble may have an amplitude of up to 1 GPa but this shock so rapidly dies down that its impact may be significant only at distances of about the initial bubble radius. However, in a concentrated bubble cluster, the combined shocks from many collapsing bubbles can cause damage at much greater distances. The feasibility of this process was supported by the experimental observations of Brunton (1967), Vyas and Preece (1976) and Shima et al. (1983).
A hypothesis that the asymmetric collapse of a bubble might generate a liquid microjet was first put forward by Kornfeld and Suvorov (1944). The first experimental and theoretical evidence of this phenomenon was provided by Naudé and Ellis (1961) and later Benjamin and Ellis (1966). Modern experimental techniques reached the stage where collapsing bubbles can be filmed at framing rates of up to 100 million frames per second: see Lauterborn (1972), Vogel et al. (1989), Field (1991), Ohl et al. (1999) and Lindau and Lauterborn (2003). Particularly impressive experiments were performed by Lindau and Lauterborn (2003). They observed the bubble jetting and shock wave emissions using high-speed cinematography of laser-produced cavitation bubbles near a rigid boundary. Experiments in which the effect of a compliant elastic boundary on the bubble collapse is investigated are reported by Brujan et al. (2001a, b). In particular, Brujan et al. observed that the interaction of a collapsing bubble with a boundary consisting of a polyacrylamide gel with 80 % water concentration could lead to, depending on conditions, bubble splitting, formation of liquid jets away from and towards the boundary, and jet-like ejection of the boundary material into the surrounding liquid.
Much work has been done on numerical modeling of bubble collapse and jet formation. Most numerical studies are based on various modifications of the boundary integral method and focus on modeling the aspherical collapse of a bubble near a rigid boundary with the formation of a liquid jet. Comparisons of measured and simulated results, for example, those made by Lauterborn and Bolle (1972) for the bubble shape at the initial stages of the collapse and Vogel et al. (1989) for liquid flow velocities during the jetting process, demonstrate good agreement.
Biomedical applications have given new impetus to studies on shock waves and microjets generated by cavitating bubbles. Kudo et al. (2009) reported that the generation of liquid microjets caused cell membrane perforation. Similar results were reported previously by Ohl et al. (2006). They investigated experimentally the interaction of a cell monolayer in the culture dish with ultrasound at high peak negative acoustic pressures (4 MPa) and observed that the high intensity ultrasound caused the detachment of cells at the focal depth. Using optical observations, they observed cell removal and showed the cells at the edge of the area became permanently porated and the surrounding cells incorporated the fluorescent marker, calcein. These observations were associated with the collapse of large cavities, which generated a strong fluid flow field. In a separate study, these cavities were assumed to be of the order of tens of micrometers as described by Le Gac et al. (2007).
Koshiyama et al. (2006, 2008) conducted a series of full atomistic molecular dynamics simulations of cell membrane models (lipid bilayers) subject to shock waves. They observed that shock impulses caused the compression and the rebound of the lipid bilayer in the course of which water molecules penetrated inside the hydrophobic core of the membrane. However, in none of these simulations, the formation of transient hydrophilic pores in the membrane occurred. It remained unclear if this was due to a rather small size of the patches considered in the simulations but in a more recent study, Koshiyama et al. (2010) have shown that such pores could form during the reorganization of a bilayer soaked with water. This effect was suggested as a possible mechanism of sonoporation, provided that shock waves lead to fast (subnanosecond) penetration of a huge quantity of water into the lipid hydrophobic region. However, it should be mentioned that their simulations did not consider the presence of gas microbubbles.
10.2.2 Hypothesized Impacts of Acoustic Phenomena on Cell Membrane and Molecular Uptake
The mechanisms involved in the sonoporation process and the cell membrane permeabilization remain poorly identified. Although no consensus has been reached, several scenarios have been hypothesized, such as the formation of pores, further stimulation of endocytotic pathways and the occurrence of membrane wounds. Elucidating the mechanisms responsible for the delivery of compounds to cells and the kinetics of permeabilization is essential for improving and controlling this therapeutic strategy.
10.2.2.1 Pore Formation
One of the earliest reports on the formation of pores on the cell membrane after ultrasound insonation dates to (1999). The study showed that applying ultrasound at 255 kHz to a suspension of HL-69 cells in presence of photosensitive drug induced cell porosity where craters were observed on the cell membrane. The authors concluded that such holes or craters are responsible for cell killing.
In 2005, the group from Bracco reported the formation of small pores in the nanometer range on MAT B III cells (2005). Using dextran markers of various sizes as drug mimics, they concluded that molecules of a diameter ranging from 11 to 37 nm crossed the membrane and entered in the cell through the formation of small pores. The group from Gent University (Geers et al. 2011) studied the transduction of cells by means of adeno-associated-virus (AAV) as a vector, which totally relies on the receptor-mediated endocytosis to successfully transducer the cells using BLM melanoma cells. They made the use of pegylated AAV, which reduces strongly the endocytosis of AAV. The cells were then tranducted using the BacMam™ 2.0 technique (GFP-reporter gene targeting the Rab5a). They observed no significant colocalization of these red-labeled vectors with the green-labeled endosomes, as no merged green and red (orange) signals could be observed inside the cells meaning that molecules ranging from 20 to 30 nm entered through pores only. These results are in contradiction with those reported earlier by the Dutch team (Meijering et al. 2009). By studying the cellular localization of fluorescent dextrans after sonoporation, they found that the smaller dextran molecules of 4.4 and 70 kDa were homogeneously distributed throughout the cytosol. This is similar to the cellular distribution found after the microinjection of dextran molecules from 3 to 70 kDa (3–8 nm) into the cytosol indicating that during sonoporation, the small dextran molecules enter cells via transient pores in the cell membrane. In contrast, dextran molecules of 155 and 500 kDa (>17 nm) were mainly localized in vesicle-like structures after sonoporation, indicating that the larger dextrans might be taken up via endocytosis. When these dextran molecules had entered via pores, a homogeneous cytosolic distribution would be expected, comparable to the distribution of these dextran molecules after microinjection. They concluded that molecules of a size less than 17 nm entered through pores while molecules larger than 17 nm entered through endocytosis pathways. Nevertheless, they did not indicate which endocytosis pathways might be involved.
Zhou et al. (2012) used controlled manipulation techniques to correlate the ultrasound-generated bubble activities with the cell membrane poration using Xenopus oocyte. They attributed the change in membrane permeability to a sub-micrometer pore caused by a local membrane rupture formed by bubble collapse or bubble compression depending on ultrasound amplitude and duration. The study was extended to investigate the sonoporation and to quantify the size and the resealing rate of the pores on a single cell using cell-attached microbubbles (2012).
Recently, Hu et al. (2013) have shown by using a cell/microbubble ratio of 1:1 that a localized perforation of the cell membrane occurs immediately upon application of a 10-cycle ultrasound pulse. It was also demonstrated that the pore creation (5.3 μm long axis diameter) was synchronized with the time of bubble collapse. This membrane disruption is transient as membrane releasing commenced within 5 s after the incidence of sonoporation, and took between 6 and 20 s to be fully restored. In contrast, large pores (12–32.6 μm) appeared following sonoporation, but did not reseal.
10.2.2.2 Endocytosis
Endocytosis pathways have been hypothesized as a mechanism of a potential plausible route for molecule incorporation into cells after sonoporation. A number of cellular reactions observed after ultrasound activation were attributed to molecular uptake, including ion exchange, hydrogen peroxide and cell intracellular calcium concentration.
Fluorescence (Lionetti et al. 2009; Meijering et al. 2009; Paula et al. 2011) and electron microscopy (Saito et al. 1999; Mehier-Humbert et al. 2005; Duvshani-Eshet et al. 2006; Yang et al. 2008; Hauser et al. 2009) studies showed an exceedingly high amount of endocytic vesicles and clathrin coated pits in sonicated cells group, while control cell sections showed single endocytic vesicles. It is believed that Ca2+ controls the uncoating of clathrin-coated pits, thus high concentrations of calcium in the cytosol are thought to trigger the uncoating of endocytic vesicles. Therefore, calcium presumably plays a crucial role in the turnover of clathrin-coated vesicles (Hauser et al. 2009).
Tran et al. (2007) investigated the implication of endocytosis routes in the sonoporation mechanisms. Ruptured-patch clamp whole-cell technique was used to measure membrane potential variations of a single cell in the presence of SonoVue microbubbles after application of ultrasound waves at 1 MHz. Microbubbles and cells were simultaneously video monitored during ultrasound exposure. The results displayed in Fig. 10.3 showed that, during sonoporation, a marked cell membrane hyperpolarization occurs at negative pressure amplitudes above 150 kPa, indicating the activation of specific ion channels, while the cell and the microbubbles remain viable. The hyperpolarization was sustained for as long as the microbubbles were in a direct contact with the cell and the ultrasound waves were transmitted. Smaller acoustic amplitudes induced only mild hyperpolarization, whereas shutting off the ultrasound brought the cell membrane potential to its resting value. However, ultrasound alone did not affect the cell membrane potential.

Fig. 10.3
Cell membrane potential after ultrasound insonation (left) and ultrasound and SonoVue microbubbles (right)
A similar hyperpolarization of the cell membrane was observed when mechanical pressure was applied to the cell through a glass probe (Fig. 10.4). The change in cell membrane potential indicates the activation of specific ion channels and depends on the quality of microbubble adhesion to the cell membrane. The use of IbTx showed that microbubbles induced a mechanical stretch activating BKCa channels. Simultaneous Ca2+ measurements indicate a slow and progressive Ca2+ increase, which is likely to be a consequence of BKCa channels opening but not a cause. In conclusion, these results demonstrate that microbubble oscillations under ultrasound activation entail the modulation of cellular function and signaling by triggering the modulation of ionic transports through the cell membrane. Cell response to the mechanical stretch caused by gentle microbubble oscillations is characterized by the opening of BKCa stretch channels and a Ca2+ flux, which might potentially trigger other cellular reactions responsible for membrane sonopermeabilization.

Fig. 10.4
 Heat-Based Tumor Ablation: Role of the Immune Response
Heat-Based Tumor Ablation: Role of the Immune Response
 MRI-Guided HIFU Methods for the Ablation of Liver and Renal Cancers
MRI-Guided HIFU Methods for the Ablation of Liver and Renal Cancers
 Design of Microbubbles for Gene/Drug Delivery
Design of Microbubbles for Gene/Drug Delivery
 Sonoporation: Applications for Cancer Therapy
Sonoporation: Applications for Cancer Therapy
 Stimulation of Bone Repair with Ultrasound
Stimulation of Bone Repair with Ultrasound
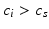 Droplets, Bubbles and Ultrasound Interactions
Droplets, Bubbles and Ultrasound Interactions




Cell membrane potential after the application of manual mechanical pressure
Related posts:
Heat-Based Tumor Ablation: Role of the Immune Response
MRI-Guided HIFU Methods for the Ablation of Liver and Renal Cancers
Design of Microbubbles for Gene/Drug Delivery
Sonoporation: Applications for Cancer Therapy
Stimulation of Bone Repair with Ultrasound
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
