CHAPTER 10 Cerebral Cortex
The surface anatomy of the cerebral hemispheres is reviewed in Chapter 9. The anatomy of the thalami and basal ganglia is reviewed in Chapter 11. Here, the focus is on the architecture of the cerebral cortex, the thalamocortical interconnections, and cerebral function. The following is a list of definitions of the prominent structures to be discussed:
Telencephalon: the most rostral portion of the brain situated above a plane directed through the anterior commissure and the velum interpositum.1 Practically, the term telencephalon equates to the cerebral hemispheres.
Cerebral cortex: the superficial layer of gray matter that extends along the surface of the two cerebral hemispheres.
Neocortex (isocortex): the pylogenetically newer portion of the cerebral cortex characterized by the presence of six predominant cell layers. The neocortex constitutes approximately 90% of the cerebral cortex (Fig 10-1).
Mesocortex: a transitional cortex interposed between the six-layered neocortex peripherally and the three-layered allocortex centrally (see later). Mesocortical tissue contains six cortical layers where it abuts the neocortex and three cortical layers where it abuts the allocortex.2 Synonyms: juxta-allocortex; paralimbic cortex.
Allocortex: the portions of the cortex phylogenetically older than the neocortex. Allocortex is characterized by the presence of three predominant layers. It comprises approximately 10% of the cerebral cortex and has two divisions: the paleocortex and the archicortex (see Fig. 10-1).
Paleocortex: the portion of the allocortex that relates to the olfactory system. It includes the olfactory bulbs, olfactory tubercles, septal region, (pre)pyriform cortex, and part of the amygdala.
Archicortex: the portion of the allocortex that relates to the hippocampal formation. It includes the hippocampus, the subicular complex, and the entorhinal cortex.
Corticoid areas: the regions of gray matter with simple, poorly differentiated cortex and no clearly discernible cortical lamination.2 The corticoid areas lie at the base of the forebrain and include the septal region deep to the paraterminal gyrus, the substantia innominata at the base of the frontal lobe, and parts of the amygdaloid complex. The corticoid regions utilize the same neurotransmitters as other cortical areas and exhibit similar interconnections.2
Limbic telencephalon: the combination of the allocortex and the corticoid areas.2
Perikaryon (plural : perikarya): the cytoplasm surrounding the nucleus of the cell (or cells).
Soma (plural : somata): the body of the cell(s), housing the nucleus and most of the protein-synthesizing apparatus.
Spine(s): dendritic protrusions that harbor excitatory synapses. Neurons that have large numbers of excitatory inputs bear large numbers of spines and are designated spiny neurons.
Cytoarchitectonics (cytoarchitecture): the systematic study of the arrangement of neuronal cell bodies within the cortex.
Myeloarchitectonics (myeloarchitecture): the systematic study of the arrangement of myelinated fibers within the cortex.1
Chemoarchitectonics (chemoarchitecture): the systematic study of the arrangement of the sources and receptors for the multiple different neurotransmitters.3–6
Radial: the direction perpendicular to both the superficial and the deep surfaces of the cerebral cortex at each point. Synonyms: vertical; perpendicular.
Tangential: the direction aligned with and coursing along the surface and laminae of the cortex at each point. Synonym: horizontal.
Unimodal: pertaining to one specific type of sensory input, such as auditory, visual, or somatosensory. Unimodal cortices receive one type of sensory input. Unimodal association areas process data received from one specific unimodal receptive cortex.
Multimodal (polymodal, heteromodal): pertaining to multiple different modalities simultaneously. Multimodal association cortices process data received from multiple different unimodal association cortices.
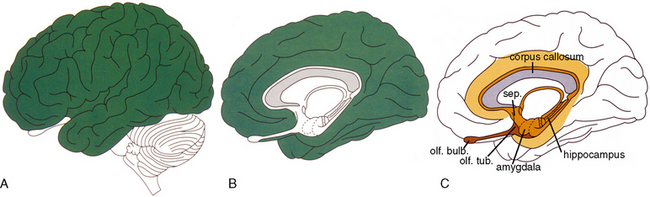
FIGURE 10-1 Human cerebral cortex. A and B, The neocortex (green) of the lateral convexity (A) and mediobasal surface (B) comprises about 90% of the cerebral surface. C, The remaining 10% is composed of the paleocortex of the olfactory system and septum (dark brown), the inner limbic ring (light brown) and the outer limbic ring (yellow orange). In humans, the outer limbic ring lies along the cingulate and parahippocampal gyri.
(From Nieuwenhuys R. The human brain: an introductory survey. Med Mundi 1994; 39:64-79.)
ANATOMY
Gross Anatomy and Telencephalization
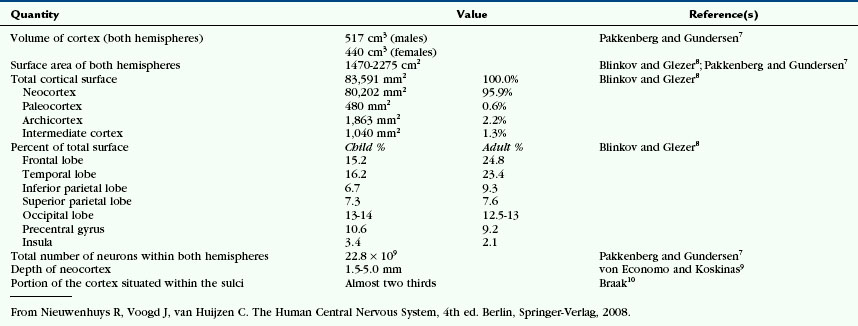
The neocortex is the phylogenetically newer portion of the cerebral cortex (Figs. 10-1 and 10-2). It comprises approximately 90% of the human cerebral cortex and is characterized by the presence of six predominant cell layers. The neocortex is especially large in humans versus other animals. On a standardized scale, the size of the neocortex is 1 in insectivores, 14.5 in prosimians, 45.5 in simians, and 156 in humans (Tables 10-1 and 10-2).1 This disproportionate overgrowth of the neocortex of the cerebral hemispheres is designated telencephalization. The newly formed telencephalic cortex provides new neural tissue for associating and processing the information entering the brain via the sensory system and for formulating sophisticated responses before initiating motor action.1
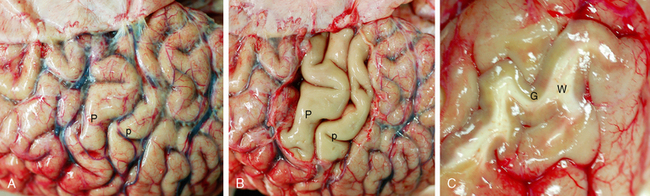
FIGURE 10-2 Cerebral neocortex. A and B, High convexity. Fresh gross specimen of the precentral (P) and postcentral (p) gyri at the level of the hand motor and sensory cortices, seen through the intact pia-arachnoid (A) and then after removal of the leptomeninges and vessels (B). C, Shearing fracture of the lateral temporal cortex displays the gray matter (G), the white matter (W), and the distinct gray-white interface.
Most of the cortex of other animals is devoted to projection areas that receive sensory data via the thalami or that help to steer motor activity.1 In these animals, the primary sensory and motor areas are separated from each other only by narrow strips of other cortex. Telencephalization expands these narrow strips into large new association cortices in the temporoparietal lobes (for sensory integration) and in the frontal lobe (for motor integration).1
Light Microscopy
Cell Types
The neocortex contains three principal cell types: (1) pyramidal cells, (2) nonpyramidal spiny neurons, and (3) nonpyramidal nonspiny neurons.11 Pyramidal cells are the most common cells of the neocortex. They are excitatory projection neurons that utilize glutamate as their neurotransmitter. Nonpyramidal spiny neurons are the next most common cell type. They are thought to be excitatory glutamatergic neurons. Nonpyramidal nonspiny neurons are the least common cell type. They are mostly inhibitory γ-aminobutyric acid (GABA)-ergic neurons.11
Pyramidal Cells
Pyramidal cells are characterized by a pyramid-shaped cell body that has its apex directed toward the surface and its base oriented tangentially, parallel to the underlying gray matter/white matter junction (Fig. 10-3).11 The apex of the pyramidal cell gives rise to a single thick apical dendrite that extends radially into the most superficial layers of the cortex. There, the apical dendrite ramifies into terminal tufts called bouquets. The apical dendrites arising from adjacent cells organize into radially oriented bundles of dendrites.
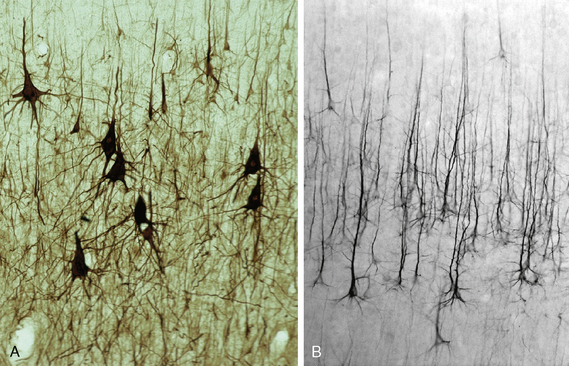
FIGURE 10-3 Pyramidal cells of the neocortex immunostained with an antibody (SMI-32) to the medium chain of the neurofilament. A, Large pyramidal cells of the motor cortex (BA 4). B, Large pyramidal cells of the parietal cortex.
(Courtesy of Dr. Patrick Hof, New York.)
The basal surface of the pyramidal cell gives rise to a single slender axon11 that extends into the underlying white matter. The axons of the pyramidal cells become myelinated shortly distal to the cell bodies and assemble into radially oriented bundles (radial fasciculi). The radial fasciculi increase in size as they descend toward the white matter and as additional axons join the bundle.1 The pyramidal cells form extensive collaterals within the cortex. Each pyramidal cell makes approximately 10,000 synapses with other cells.
Nonpyramidal Spiny Cells
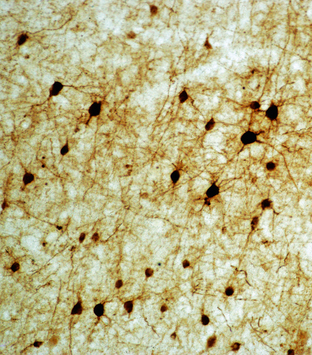
Nonpyramidal spiny cells (synonyms: spiny granule cells; spiny stellate neurons) are small, multipolar cells that give rise to limited numbers of primary dendrites (Fig. 10-4). These dendrites fan outward in multiple directions and are densely covered in spines. The axons arising from these cells branch outward, predominantly radially.11
Cortical Architecture
The architecture of the cortex shows distinct tangential and radial elements.
Lamination
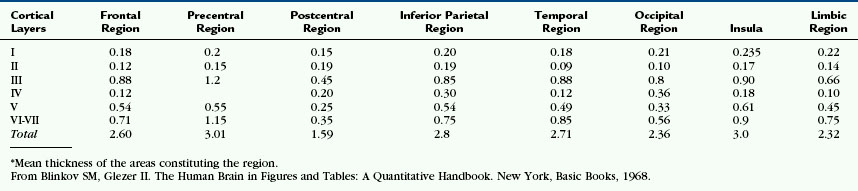
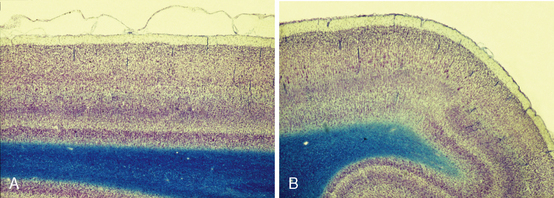
The neocortex consists of six tangential zones or layers. Each layer is characterized by the number, type, and arrangement of the cell bodies (perikarya) within it and by the organization of the myelinated fibers that course through it (see Tables 10-2 and 10-3).1 From superficial to deep, these layers are numbered I to VI and are designated by the cell type once thought to be predominant within each layer (Figs. 10-5 to 10-7).

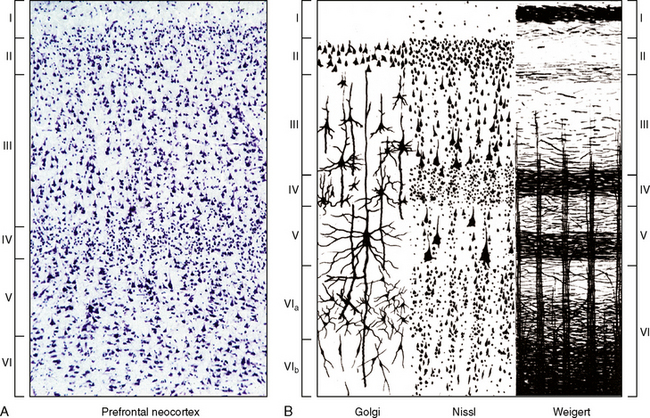
FIGURE 10-5 Lamination of the neocortex: layers I to VI. A, Histologic section of the six-layered human neocortex. Nissl stain for neurons. B, Diagrammatic representation of the cytoarchitecture and myeloarchitecture of the six layers of the neocortex. Column 1. The Golgi stain impregnates the entire neuron, showing the location and full extent of the cells in each layer. Column 2. The Nissl stain demonstrates the cell bodies, showing the location and lamination of the somata of the cells. Column 3. The Weigert stain for myelinated nerve fibers demonstrates the radial (columnar) and horizontal (laminar) arrangement of nerve fibers in each layer. The myelinated plexi define sublayers within each of the six principal layers.
(A, Courtesy Dr. Patrick Hof, New York. B, From Carpenter MB, Sutin J. Human Neuroanatomy 8th Ed. 1983 Baltimore, Williams & Wilkins).
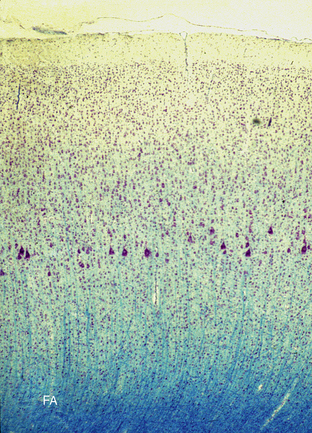
FIGURE 10-6 Cortical columns in the primary motor cortex (Klüver-Barrera stain) (non-human primate). The cerebral cortex displays distinct radial, columnar organization in addition to the laminar architecture. Note the large pyramidal (Betz) cells in the internal pyramidal cell layer V.
Overview of the Cortical Layers
Layer I. The molecular layer may be thought of as the primordial input layer, because it receives axons from many early, highly conserved structures. These include the noradrenergic locus ceruleus, the serotonergic dorsal raphe nucleus, the dopaminergic ventral tegmental area, and cholinergic cells within the nucleus accumbens septi. None of these inputs is restricted to this layer. Layer I also receives corticocortical synapses from most other cell layers and projections from the anterior and intralaminar thalamic nuclei.
Layer II. The external granular cell layer is the most superficial layer of corticocortical neurons. Because these cells lie closest to layer I, they have the shortest apical dendrites and look “granular.” That appearance gave rise to the misnomer “external granular layer.” Layer II contains many inhibitory interneurons, adding to its nonpyramidal appearance.
Layer III. The external pyramidal layer is composed of predominantly corticocortical pyramidal neurons and inhibitory interneurons.
Layer IV. The internal granular cell layer can be thought of as a major input layer of the cortex. It receives substantial thalamic inputs from phylogenetically more recent thalamic nuclei, such as the ventral posterolateral, ventral posteromedial, lateral geniculate, and medial geniculate nuclei. It also contains inhibitory interneurons. In cortical areas that primarily have “output” function, layer IV is very thin.
Layer V. The internal pyramidal layer is a major output layer of the cortex. It contains pyramidal neurons that project both cortically and subcortically.
Layer VI. The multiform layer contains neurons that project subcortically and corticocortically as well as interneurons.
Cortical Columns
The cortical neurons organize into radial columns that extend through all six cortical layers, superficial to deep (see Fig. 10-6). These radially oriented cortical columns appear to be the fundamental units, or modules, for cortical function.
Within the primary sensory areas of the neocortex (i.e., the auditory, visual, and somatosensory cortices), the neurons are arranged as small radial columns that surround a radially oriented thalamic afferent fiber. In other areas of the neocortex, the neurons form radial columns organized around corticocortical afferents rather than thalamic inputs.1 Within the visual cortex, for example, the radial organization establishes orientation columns approximately 300 to 500 μm in diameter, ocular dominance columns approximately 500 μm in width, and ellipsoidal “blobs” 150 to 200 μm in width. Within the motor cortex, radial cortical motor columns approximately 1 mm in diameter appear to control the contraction of specific muscles.1 (See the later sections on myeloarchitecture, classification by thalamic connections, cortical afferents to the neocortex, and function.)
Parcellation of the Cortex
From area to area across the cerebral hemisphere, the cortex shows differences in the relative thickness of its gray matter; the thickness and cell density of each cortical layer; the nature and arrangement of the neuronal perikarya within the layers; the packing density and laminar arrangement of the myelinated fibers; and the density and laminar distribution of receptors for multiple neurotransmitters (see Figs. 10-5 to 10-7).1 These differences are used to partition the cortex into cyto/myelo/chemoarchitectonic areas that correspond to the different functions of each portion of cortex, at least in part.9
Korbinian Brodmann12 parcellated the human cortex into 52 cytoarchitectonic areas now designated the Brodmann areas (BA). At present, this Brodmann map (Fig. 10-8) is the most widely used system for identifying functional areas, locations of pathologic processes on routine neuroimages, and sites of activation on functional MR images. Nieuwenhuys and colleagues estimate that there may actually be 150 juxtaposed structural (and potentially functional) areas present within the human cortex.1 Other authors have described up to 200 separate cortical areas.9,13
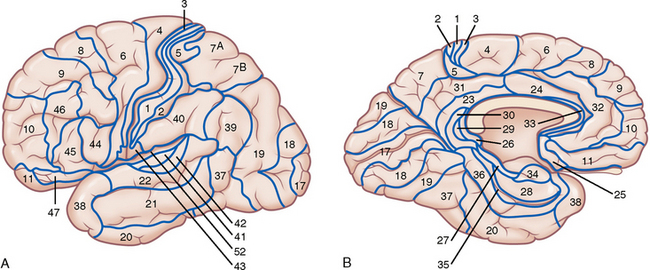
Gross Organization of the Cortex with Brodmann Designations
Frontal Lobe
Primary Motor Cortex (MI) (BA 4)
The primary motor cortex (MI) is a long “triangular” region situated along the length of the precentral gyrus (see Fig. 10-8). On the lateral surface, superiorly, MI occupies the full anteroposterior extent of the upper precentral gyrus. Inferiorly, MI tapers progressively, so its thin lower portion is confined to the posterior face of the precentral gyrus within the central sulcus. On the medial surface, MI occupies most of the paracentral lobule. Histologically, the primary motor cortex conforms to BA 4. In BA 4, the internal granule cell layer (IV) is nearly absent. The internal pyramidal cell layer V is thick and characteristically displays clusters of very large pyramidal cell bodies—the Betz cells—whose axons extend into the corticospinal and corticobulbar tracts.11
Premotor Cortex (PM) (BA 6)
The premotor cortex occupies a large portion of the frontal lobe immediately anterior to BA 4 (see Fig. 10-8).11 On the convexity, BA 6 lateral extends over the frontal convexity and corresponds to the premotor (PM) area. This is subdivided into dorsal (d) and ventral (v) portions. The dorsal premotor area (PMd) receives input from the dorsolateral prefrontal region, whereas the ventral premotor area (PMv) receives input from the ventrolateral prefrontal region.11 On the medial surface, BA 6 lies anterior to the paracentral lobule and extends from the superior margin of the hemisphere peripherally to the cingulate sulcus (BA 24) below.
Prefrontal Cortex (PF) (BA 9, BA 46, and BA 45)
The prefrontal cortex is also subdivided into dorsal and ventral portions (PFd and PFv) (see Fig. 10-8). PFd largely corresponds to BA 9 (and perhaps superior BA 46). PFv largely corresponds to inferior BA 46 and BA 45.11 The medial prefrontal cortex includes BA 32 and 25. This region is similar to the anterior cingulate cortex (BA 24), so the two are often considered to be a single complex.1
Frontal Pole (FP) (BA 10)
BA 10 lies along the convexity and the medial surface of the superior frontal gyrus at the frontal pole (see Fig. 10-8). The human frontopolar cortex is specifically devoted to complex cognitive functions, such as integrating the outcomes of two or more separate cognitive operations directed toward a higher behavioral goal.1
Broca’s Area (BA 44 and Part of BA 45)
In the dominant hemisphere, Broca’s motor speech area is classically considered to lie within the inferior frontal gyrus at BA 44 and the adjacent portion of BA 45 (see Figs. 10-8 and 10-9). Some authorities now contest that localization and suggest that the true Broca motor speech area lies, instead, in the anterior lobule of the insula just deep to the inferior frontal gyrus.11,14
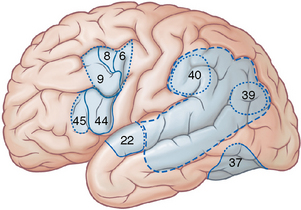
FIGURE 10-9 Language areas of the brain and frontal eye fields. Broca’s motor speech area lies in the inferior frontal gyrus at the pars opercularis (BA 44) and the adjacent portion of the pars triangularis (BA 45). Wernicke’s receptive area for speech has uncertain borders and may be highly individual. It appears to be included within the supramarginal (BA 40) and angular (BA 39) gyri of the inferior parietal lobule. BA 22 of the superior temporal gyrus is related to auditory processing of speech. BA 37 may have visuoauditory functions for speech recognition. The frontal eye field includes portions of BA 6, 8, and 9.
(Modified from Standring S [ed]. Cerebral hemisphere. In Gray’s Anatomy: The Anatomical Basis of Clinical Practice, 40th ed. Philadelphia: Churchill Livingstone Elsevier, 2008.)
Supplementary Motor Area (MII) (SMA) (BA 6aα)
The SMA lies on the medial surface of the frontal lobe (Fig. 10-10). Zilles has localized the SMA to the portion of the medial cerebral cortex situated between two specific landmarks along the Talairach-Tournoux baseline. The Talairach-Tournoux baseline is the line drawn from the top of the anterior commissure (AC) to the bottom of the posterior commissure (PC). The anterior landmark, VAC (vertical at the anterior commissure), is the line raised perpendicular to the Talairach-Tournoux baseline at the anterior commissure. The posterior landmark, VPC (vertical at the posterior commissure), is the line raised perpendicular to the Talairach-Tournoux baseline at the posterior commissure.11,15 The supplementary motor area serves for learning and generating sequences of actions, for selecting the side to use for unilateral motor action, and for coordinating bimanual action and posture.16
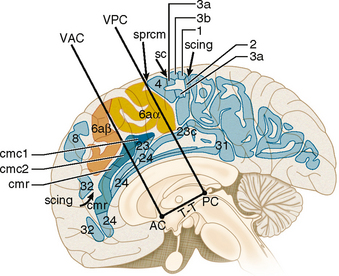
FIGURE 10-10 The supplementary motor area (SMA) and cingulate motor area. The supplementary motor area (BA 6aα) lies on the medial surface at the paracentral lobule between VAC and VPC (the verticals erected to the Talairach-Tournoux (T-T) baseline at the anterior commissure (VAC) and posterior commissure (VPC). The pre-SMA (negative SMA) (BA 6aβ) lies just anterior to the SMA. The cingulate motor cortex includes a rostral zone (cmr) that lies entirely rostral to VAC and a caudal zone (cmc) that flanks VAC but lies entirely rostral to VPC. Other labeled structures include the central sulus (sc), cingulate sulcus (scing), medial precentral sulcus (sprcm), and the numbered Brodmann areas.
(From Zilles K, Schlag G, Geyer S, et al. Anatomy and transmitter receptors of the supplementary motor areas in the human and nonhuman primate brain. Adv Neurol 1996; 70:29-43.)
Presupplementary Motor Area (pre-SMA) (BA 6aβ)
The pre-SMA lies on the medial surface of the frontal lobe anterior to the SMA (see Fig. 10-10). It is directly involved in motor inhibition. The pre-SMA is involved in selecting appropriate motor responses by suppressing automatic responses to environmental stimuli and stopping previously planned actions as new data indicate a need for change.17
Cingulate Gyrus
The cingulate cortex (see Fig. 10-10) contains multiple co-curvilinear regions that extend from beneath the genu of the corpus callosum (CC) (subgenual cingulate cortex), around and anterior to the CC (pregenual or perigenual cingulate cortex), then above the CC (supragenual cingulate cortex) to behind the splenium (retrosplenial cingulate cortex) (see Figs. 10-8, 10-10, and 10-11). These regions include BA 25 in the subgenual cingulate cortex; BA 33, 24 a/b/c, and 32 in the pregenual and supragenual cingulate cortex; caudal BA 32′ and caudal BA 24′ in the dorsal anterior cingulate cortex; BA 23 and 31 in the posterior cingulate cortex, and BA 29 and 30 in the retrosplenial cingulate cortex. As used in the neuropsychological and psychiatric literature (see Figs. 10-10 and 10-11), the term rostral anterior cingulate cortex conforms to the pregenual CC and includes BA 32 and inferior portions of BA 24. The subgenual anterior cingulate cortex lies inferior to the genu and includes BA 25 and caudal portions of BA 32 and 24. The dorsal anterior cingulate cortex includes caudal BA 24′ and 32′ and the cingulate motor area. The anterior cingulate region also contains affective and cognitive subdivisions.18
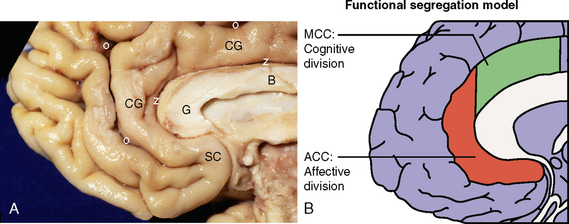
FIGURE 10-11 Cingulate gyrus. A, Medial brain surface. The cingulate gyrus (CG) extends around the body (B) and genu (G) of the corpus callosum between the callosal sulcus (z) centrally and the cingulate sulcus (o) superficially. The subgenual portion of the cingulate gyrus contains the subcallosal area (SC). B, On anatomic and physiologic grounds, the rostral cingulate cortex consists of functionally distinct regions: a rostroventral affective division (ACC or ventral ACC), and a dorsal cognitive division (MCC).
(B, Redrawn from Shackman AJ, Salomons TV, Slagter HA, et al. The integration of negative affect, pain, and cognitive control in the cingulate cortex. Nat Rev Neurosci 2011; 12:154-167; and based on data from Bush G, Luu P, Posner MI. Cognitive and emotional influences in anterior cingulate cortex. Trends Cogn Sci 2000; 4:215-222.)
Parietal Lobe
Somatosensory Cortex (S1) (BA 3a, BA 3b, BA 1, and BA 2)
S1 lies along the convexity surface and superior medial surface of the postcentral gyrus (see Fig. 10-8). At both sites, BA 3b, BA 1, and BA 2 form long vertical strips of tissue along the length of the postcentral gyrus. In order from anterior to posterior: BA 3b is buried within the central sulcus along the anterior face of the postcentral gyrus; BA 1 lies behind it along the posterior lip of the central sulcus; and BA 2 lies farther back along the crown of the postcentral gyrus. There may be serial, hierarchical processing of data from BA 3b through BA 1 to BA 2.11 Parietal region BA 3a, which abuts directly on MI, is usually considered with the motor cortex.1 Histologically, BA 1 through BA 3 are distinguished by the thinness of their cortices overall and by their especially thick, very compact layer IV.
Secondary Somatosensory Cortex (SII) (BA 5 and parts of BA 40 and BA 43)
SII lies along the upper margin of the sylvian fissure in the medial parietal operculum, just behind the central sulcus.11 An additional parietal ventral area (PV), along the medial aspect of the parietal operculum, is also considered to be part of the somatosensory cortex.1
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


