Normal Abdominal and Pelvic Anatomy
Dennis M. Balfe
Brett Gratz
Christine Peterson
More than ever, a successful radiologist must be a practical applied anatomist. To become effective, the radiologist has had to assimilate the standard anatomic information provided by classical dissections. That information is, however, to an increasing extent, inadequate to explain the dynamic anatomic changes observed in pathologic conditions. Radiologists continue to enrich and modify standard conceptions of important structures in the abdomen and pelvis. Examples of the importance of precise anatomic observations abound in the literature: tiny extensions of the pancreatic ductules within a mass can be the sole differential point in diagnosing tumefactive pancreatitis; thin, nearly invisible, fascial planes determine the distribution of fluid collections within the retroperitoneum. The importance of a thorough understanding of human anatomy has grown exponentially as our ability to image it has been refined.
One of the first principles of radiologic practice is that one must first recognize normal structures, including all of their normal variations, in order to confidently diagnose what is abnormal. Now that imaging technology has advanced to the point that subtle anatomic details can be resolved and displayed, radiologists must learn to recognize them and to understand their behavior in health and disease.
The most pervasive single change to occur in the past decade has been the ability to render volumetric data sets in any user-defined anatomic plane, in “rubber-sheet” curved planes, or as images with a component of three- dimensional information content. Although this ability has been available as a laboratory tool for 20 years, the ability to capture a crisp data set made of isotropic voxels, coupled with the introduction of user-friendly software tools, has made three-dimensional (3D) imaging a clinical reality. Accordingly, although many examples of normal abdominal and pelvic anatomy are still well depicted on axial sections, there are many anatomic regions better displayed in other planes (149) or using volumetric display. Accordingly, this chapter will incorporate multiplanar and volumetric renderings whenever appropriate. The internal structure of abdominal organs will be described, but only briefly; the reader is referred to more detailed discussions of the anatomy of each organ in the chapter dealing with that specific system. In all cases, particular emphasis will be placed on those anatomic details that have practical applications to daily imaging procedures and to those regions in which pathologic processes are likely to occur.
ABDOMINAL ANATOMY
Abdominal Wall
The rectus abdominis muscles compose the anterior aspect of the abdominal wall (47) (Fig. 10-1). They attach to the front of the xiphoid process and to costal cartilages 5 to 7. Extending inferiorly as flat, relatively broad structures, they attach to the pubic symphysis. Above the umbilicus, the rectus muscles are surrounded by a strong sheath formed by the aponeuroses of the three anterolateral muscles (99): the anterior layer is formed by fibers of the external oblique and by a portion of the fibers of the internal oblique. The posterior layer is formed chiefly by fibers from the transverses abdominis muscle, as well as some from the internal oblique. However, about 2 cm below the umbilicus, the posterior portion of the sheath disappears, and fibers of all
three anterolateral muscle groups pass anterior to the rectus muscle (the zone of anatomic transition is called the arcuate line). This arrangement has clinical significance in that rectus sheath hematomas that occur in the upper abdomen are well confined inside the rectus sheath. Inferior to the arcuate line, however, they can escape into the easily expandable transversalis fascia, and can dissect posteriorly (into the space of Retzius), across the midline, or laterally into the flank (Fig. 10-2).
three anterolateral muscle groups pass anterior to the rectus muscle (the zone of anatomic transition is called the arcuate line). This arrangement has clinical significance in that rectus sheath hematomas that occur in the upper abdomen are well confined inside the rectus sheath. Inferior to the arcuate line, however, they can escape into the easily expandable transversalis fascia, and can dissect posteriorly (into the space of Retzius), across the midline, or laterally into the flank (Fig. 10-2).
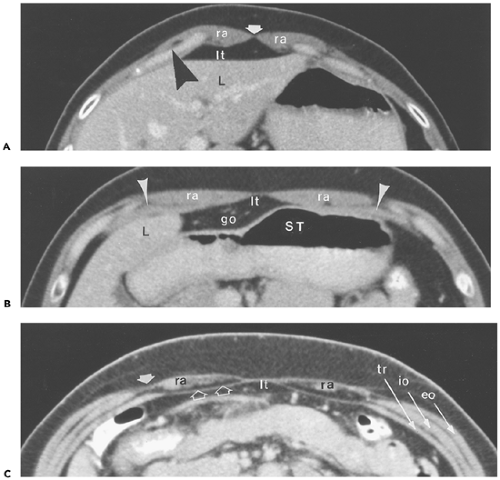 Figure 10-1 Abdominal wall: rectus muscle and sheath. A: Section obtained immediately caudal to the xiphoid shows the paired rectus abdominis muscles (ra), which narrow medially to attach to the linea alba (arrow). They attach laterally to the costal cartilages (arrowheads) of the fifth through seventh ribs. The fat immediately posterior to them lies in the root of the ligamentum teres (lt). L, left hepatic lobe. B: Section obtained 5 cm caudal to (A). The rectus muscles (ra) have thinned and broadened. Laterally, they lie on the surface of the transversus abdominis muscle (arrowheads); medially, the fat in the root of the ligamentum teres (lt) apposes their posterior surfaces. Posterior to that, on the right, is a portion of the greater omentum (go); on the left is the body of the stomach (ST). L, liver. C: In this patient, fatty infiltration of both rectus abdominis muscles (ra) allows observation of the rectus sheath. At this level (above the arcuate line), fibers from the transversus abdominis muscle (tr) and, variably, from the internal oblique muscle (io) pass posterior to the rectus abdominis muscle to form the aponeurotic posterior rectus sheath (open arrows). Fibers of the external oblique (eo) and internal oblique muscles blend together (arrow) to course over the rectus abdominis muscle, forming the anterior rectus sheath. lt, fat in the root of the ligamentum teres. |
Superficial to the rectus muscles, within the subcutaneous fat, are the superficial epigastric veins, which are generally largest on sections through the pelvis (Fig. 10-3). The inferior
epigastric vessels course between the belly of the rectus muscle and the posterior rectus sheath; on sections below the arcuate line, they lie between the rectus muscle and the transversalis fascia. On consecutively inferior sections, the inferior epigastric vessels course laterally within the lateral umbilical fold, which marks the medial aspect of the deep inguinal ring. The vessels also form the lateral boundary of the inguinal triangle (the Hasselbach triangle), the medial boundary of which is the edge of the rectus muscle, and the inferior boundary of which is the pubic bone. Coronal and sagittal images best depict the origin and course of the inferior epigastric artery and vein (see Fig. 10-3).
epigastric vessels course between the belly of the rectus muscle and the posterior rectus sheath; on sections below the arcuate line, they lie between the rectus muscle and the transversalis fascia. On consecutively inferior sections, the inferior epigastric vessels course laterally within the lateral umbilical fold, which marks the medial aspect of the deep inguinal ring. The vessels also form the lateral boundary of the inguinal triangle (the Hasselbach triangle), the medial boundary of which is the edge of the rectus muscle, and the inferior boundary of which is the pubic bone. Coronal and sagittal images best depict the origin and course of the inferior epigastric artery and vein (see Fig. 10-3).
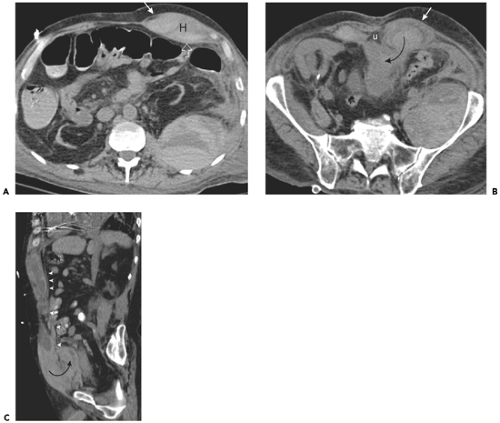 Figure 10-2 Rectus sheath. A: Section obtained above the arcuate line in a patient with left rectus sheath hematoma (H). Note that the hematoma is well confined between the aponeurotic fibers of the external oblique (arrow) anteriorly and the transversus abdominis (open arrow) posteriorly. B: Section obtained below the arcuate line shows that only the anterior aponeurosis (arrow) persists. Posteriorly, the hematoma is unconfined, and extends (curved arrow) into the extraperitoneal space. Note triangular fat surrounding the urachus (U). C: Sagittal image in another patient with extensive rectus sheath hematoma shows that posterior layer of the rectus sheath (arrowheads) confines the hematoma to the level of the arcuate line. Caudal to this, the hematoma is free to extend (curved arrow) into the prevesical fat. |
The anterolateral abdominal wall is made up of three paired muscles: from superficial to deep, they are the external oblique, internal oblique, and transverses abdominis. As noted above, the medial aspect of all three muscles is an aponeurosis, which contributes to the formation of the rectus sheath. The lower part of the external oblique aponeurosis forms the inguinal ligament. Magnetic resonance imaging (MRI) has been used effectively to detect strains of the anterolateral musculature (23).
The major muscles of the posterior abdominal wall are, medially, the erector spinae muscles and, laterally the latissimus dorsi (Fig. 10-4). The erector spinae (or sacrospinalis) muscle is really a group of three muscles that cannot be distinguished using standard imaging techniques: from lateral to medial, they are the iliocostalis, the longissimus, and the spinalis. Combined, these muscles are narrow at sacral levels but broad in the thoracolumbar region (54).
The posterolateral surface of the external oblique muscle is covered by the latissimus dorsi, except inferiorly, where the two muscles attach in separate locations to the iliac
crest. The external oblique muscle attaches anteriorly, whereas the tendon of the latissimus dorsi passes posteriorly. This exposes a small triangle of internal oblique musculature just above the iliac crest. This inferior lumbar triangle (also known as the Petit triangle) (Fig. 10-5) is the site of spontaneous lumbar hernias (11). This area is important to scrutinize in patients with high-speed vehicular trauma, particularly in patients restrained by seat belts (70) (Fig. 10-6).
crest. The external oblique muscle attaches anteriorly, whereas the tendon of the latissimus dorsi passes posteriorly. This exposes a small triangle of internal oblique musculature just above the iliac crest. This inferior lumbar triangle (also known as the Petit triangle) (Fig. 10-5) is the site of spontaneous lumbar hernias (11). This area is important to scrutinize in patients with high-speed vehicular trauma, particularly in patients restrained by seat belts (70) (Fig. 10-6).
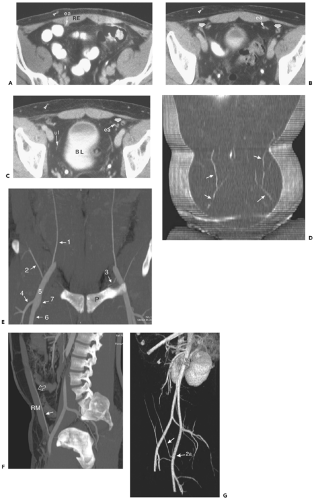 Figure 10-3 Superficial epigastric vessels. A: Section through the pelvic inlet shows the superficial epigastric vein (arrowhead) within the subcutaneous fat anterior to the rectus abdominis muscle (RE). This vein is part of an extensive network that connects to the thoracoepigastric system, the inferior epigastric system, and the paraumbilical venous plexus. Just deep to the rectus abdominis muscle is the epigastric artery (ea) and vein. B: The epigastric vessels (ea) 16 mm inferior to (A) lie close to the vasa deferentia (arrows) in men (the round ligament occupies this position in women). Arrowhead, superficial epigastric vein. C: The epigastric vessels (ea) 16 mm inferior to (B), near their site of origin from the external iliac artery and vein, cross the vasa deferentia (arrow). Nearer the midline is the medial umbilical ligament (ul), the obliterated remnant of the umbilical artery, coursing anterior to the bladder (BL). Arrowhead, superficial epigastric vein. D: Coronal view of the anterior abdominal wall shows the serpentine course of the superficial epigastric veins (arrows). E: Coronal thick-slab MIP showing the relationship of the inferior epigastric arteries (1) with major superficial branches of the external iliac artery. The inferior epigastric artery arises just above the inguinal ligament, whereas the superficial circumflex iliac artery (2) usually arises from the common femoral artery just caudal to the inguinal ligament. The proximal inferior epigastric artery gives rise to the pubic artery (3), which courses caudally towards the pubis (P); inferiorly, it forms an anastamosis with the obturator artery. The common femoral artery (5) bifurcates into the superficial femoral (6) and deep femoral (7) arteries. The latter gives rise to a large lateral branch, the lateral circumflex femoral artery (4). F: Sagittal thick-slab maximum-intensity projections shows the course of the inferior epigastric artery (arrow) on the posterior border of the rectus muscle (RM); superiorly, it pierces the transversalis fascia (open arrow) just below the arcuate line and ascends between the rectus muscle and its sheath. G: Oblique perspective volume rendered image shows the origin of the left inferior epigastric artery (arrow) from the external iliac artery (ea). |
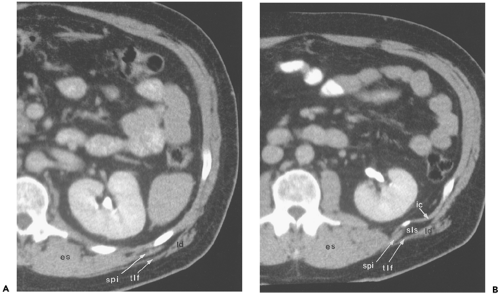 Figure 10-4 Posterior abdominal wall. A: Section at the level of the renal hilus shows the bulky erector spinae muscles groups (es) adjacent to the vertebral transverse process. Overlying the ribs on the posterolateral surface of the body are the serratus posterior inferior (spi) and latissimus dorsi (ld) muscles. The latter gives rise to a tough fascial layer, the thoracolumbar fascia (tlf). B: Section obtained 16 mm caudal to (A). The superior lumbar space (sls), here containing a small amount of herniated fat, lies between the intercostal muscles (ic) and the latissimus dorsi muscle (ld) and thoracolumbar fascia (tlf) just lateral to the serratus posterior inferior (spi) muscle band. es, Erector spinae. |
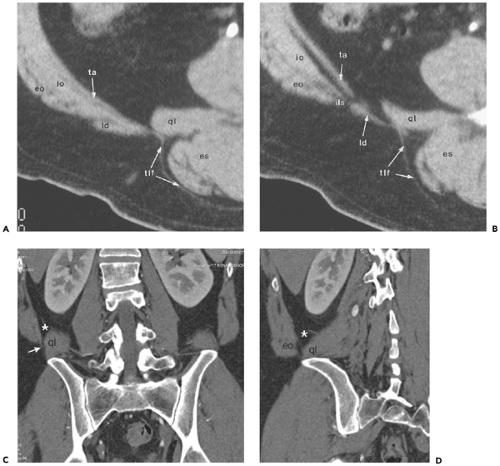 Figure 10-5 Inferior lumbar triangle (the Petit triangle): normal anatomy. A: Section through the quadratus lumborum muscle (ql) shows the band-like latissimus dorsi muscle (ld) covering the posterior aspect of the posterior abdominal wall, comprised of the transversus abdominis (ta), internal oblique (io), and external oblique (eo) muscles. The thoracolumbar fascia (tlf), an extension of the latissimus dorsi muscle, extends posteromedially to cover the surface of the quadratus lumborum and erector spinae (es) muscles. B: Section obtained 7 mm caudal to (A) shows that the latissimus dorsi (ld) muscle has passed posteromedially with respect to the posterolateral abdominal musculature [transversus abdominis (ta), internal oblique (io), and external oblique (eo) muscles] to create a defect, the inferior lumbar space (ils) through which lumbar hernias can protrude. ql, Quadratus lumborum muscle; es, erector spinae; tlf, thoracolumbar fascia. C: Coronal reformation in another patient shows the fat-containing inferior lumbar triangle (asterisk) between the quadratus lumborum muscle (ql) medially and the thin fibers of the transversus abdominis muscle (arrow) laterally. D: Oblique coronal reformation highlights the fat-containing space (asterisk) between the abdominal wall musculature (chiefly composed of the external oblique muscle (eo) and the quadratus lumborum muscle (ql). |
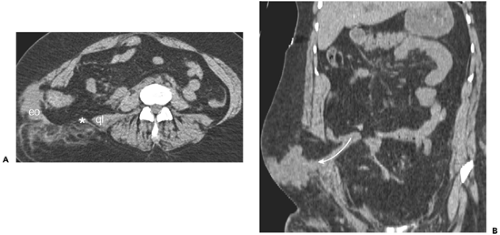 Figure 10-6 Traumatic lumbar hernia. A: Axial section obtained above the iliac crest in a patient involved in high-speed motor vehicle collision while wearing a lap belt. There is marked separation between the lateral abdominal wall musculature (eo) and the latissimus dorsi muscle (asterisk), lying just lateral to the quadratus lumborum (ql). B: Coronal section in the same patient shows herniation of small intestine (curved arrow) through the traumatic hernia. |
There is, likewise, a relative point of weakness in the aponeuroses of the transversus abdominis and internal oblique muscles just lateral to the rectus abdominis muscle near the level of the arcuate line. This is the classic site for a Spigelian hernia (126). These hernias, when small, are frequently clinically inapparent, because they are confined deep to the strong external oblique muscle (Fig. 10-7).
The inguinal canal is the major structure passing out of the abdomen, through the lateral wall musculature. In men, the canal contains the spermatic cord; in women, the round ligament. The deep inguinal ring is a slit-like opening in the transversalis fascia. As its contents pass through the inguinal canal, they are covered by fibers of the internal oblique muscle, which, inferiorly, form the cremaster muscle. Finally, they pass inferomedially through the aponeurotic part of the external oblique at the superficial inguinal ring. Indirect inguinal hernias (which are by far the more common) essentially follow the path of the inguinal contents and therefore begin just lateral to the inferior epigastric vessels (Fig. 10-8). Direct inguinal hernias protrude through defects in the transversalis fascia that comprises the floor of the inguinal canal; they occur medial to the inferior epigastric vessels (77,150).
Computed tomography (CT) is useful in depicting abdominal wall hernias, which, in many cases, are difficult to evaluate clinically because of obesity or scarring from previous surgery. Scans performed with the patient in a lateral decubitus position, or during a Valsalva maneuver, may demonstrate hernias that are not well depicted during standard imaging techniques (1,33,52,65).
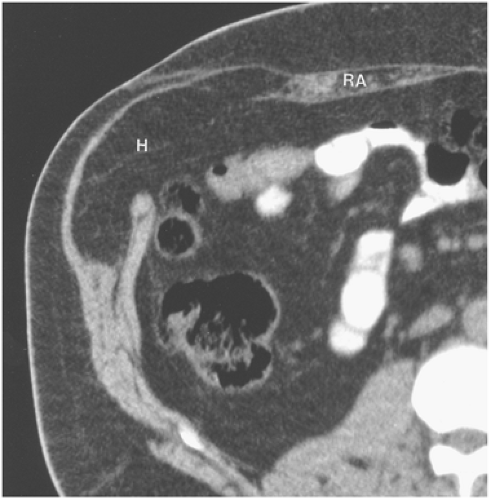 Figure 10-7 Spigelian hernia. Transverse computed tomography section obtained through the midabdomen shows herniation of greater omentum (H) through a defect immediately lateral to the rectus abdominis muscle (RA). This is the classic location for a Spigelian hernia. |
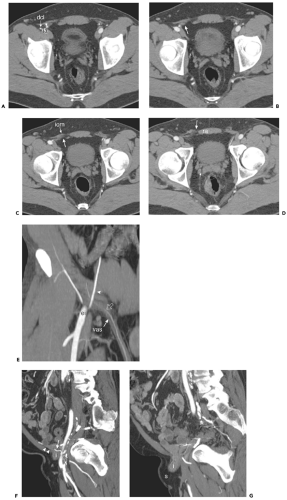 Figure 10-8 The inguinal canal. A: Axial image obtained in a normal patient shows the inferior epigastric artery (arrowhead) arising from the external iliac (ia) at the level of the internal inguinal ring. The deep circumflex iliac artery (dci) arises at nearly the same level. B: Axial image 5 mm inferior to (A) shows the spermatic cord (arrow) entering the right inguinal canal. C: Axial image 5 mm inferior to (B) shows the right spermatic cord (arrow) coursing medially within the inguinal canal, behind fibers of the internal oblique muscle (iom). D: Axial image 5 mm inferior to (C) shows the spermatic cord (arrow) emerging from the external inguinal ring to lie just lateral to the rectus abdominis muscle (ra). E: Oblique coronal reformatted image shows the inferior epigastric artery (arrowhead) arising from the external iliac (ei); the inferior epigastric gives rise to the cremasteric artery (open arrow) which accompanies the vas deferens (vas) into the inguinal canal. F: Sagittal image in another subject with a direct inguinal hernia shows the inferior epigastric artery (arrow) which marks the lateral boundary of the Hasselbach triangle. The origin of the inferior epigastric artery and its pubic branch (open arrow) marks the position of the internal inguinal ring. In this patient, an intestinal loop (i) protrudes through a defect in the transversalis fascia (arrowheads) to enter the inguinal canal. G: Sagittal image obtained 10 mm medial to (E) shows intestinal loop (i) extending toward the fat within the scrotum (s). |
Diaphragm, Crura, and Arcuate Ligaments
The diaphragm is a large, dome-shaped muscle that incompletely divides the thorax from the abdomen. Its fibers take origin from the sternum anteriorly, from the medial surfaces of the lower ribs anterolaterally, and from the upper lumbar vertebral bodies posteriorly. They insert superomedially on an aponeurosis (the central tendon), the thinnest portion of the diaphragm, which is shaped like an inverted “V” on transaxial sections. The apex of the central tendon lies just anterior to the inferior vena cava; its limbs descend posterolaterally to enclose the vena cava on the right and pass anterior to the esophageal hiatus on the left. It is through the esophageal hiatus that hiatal hernias (as well as ascites, or pancreatic fluid collections) can extend into the posterior mediastinum (84) (Fig. 10-9).
The cross-sectional appearance of the relatively short anterior fibers of the diaphragm depends on the patient’s body habitus and resultant position of the middle leaflet of the diaphragm relative to the sternum. Normally, the central tendon lies 2 to 3 cm cephalic to the xiphoid process, so that cross-sectional images of the diaphragm show a thin soft tissue stripe crossing roughly parallel to the anterior body wall. A more caudally positioned central tendon can produce a confusing image (40), resulting in an anterior pseudomass close to the xiphoid process (Fig. 10-10). In this situation, anterior diaphragmatic fibers course anterior and lateral to their origin on costal cartilage and enclose abdominal fat, forming an arch anterior to liver and heart. This orientation produces fan-like soft tissue stripes projecting from the base of the heart to the anterior body wall. If the central tendon and xiphoid are at the same level, most of the anterior diaphragm will be imaged on a single slice and will appear as a very broad soft tissue band. This portion of the diaphragm is not optimally imaged in the axial plane; these confusing images can easily be resolved using images reformatted in sagittal or coronal planes.
In most patients, the lateral and posterior portions of the diaphragm are perpendicular to the plane of axial section and are displayed as thin soft tissue stripes separating lung parenchyma from abdominal fat. Suspended inspiration, particularly in patients who perform a Valsalva maneuver, allows the muscular slips of the diaphragm to relax and become folded near their costal insertions. They are then displayed as discrete, thick, sometimes nodular soft tissue densities, which may indent the liver, stomach, colon, or spleen, forming accessory fissures or pseudotumors. This is commonly observed in elderly individuals (118) (Figs. 10-11 and 10-12). These infoldings can be distinguished from pathologic nodular densities by noting their continuity with the diaphragm and their separation from abdominal solid and hollow viscera by subdiaphragmatic fat. In problematic cases, repeat imaging in expiration will confirm their identity as diaphragmatic structures (5,117).
The lumbar, or posterior, portion of the diaphragm arises from the crura and the medial and lateral arcuate ligaments. The right and left diaphragmatic crura take origin from the anterolateral surface of the first three right and first two left lumbar vertebral bodies. They unite anteriorly
to form the median arcuate ligament, surrounding the aorta immediately cephalic to the celiac trunk (110). This ligament can be imaged as a thin soft tissue stripe crossing the aorta; in some cases, it can produce deformity of the celiac trunk (Fig. 10-13) (134). The right crus is larger and originates lower than the left; it extends to the left of midline. As the crura continue upward, the right crus divides to enclose the esophagus within the esophageal hiatus. As the esophagus enters the stomach, both structures are relatively firmly tethered to the fissure for the ligamantum venosum
by the gastrohepatic ligament. As a result, a portion of the stomach wall runs in a transverse plane, and therefore appears thicker than the rest of the stomach on transverse sections (28). This “pseudomass” has been noted in 25% to 30% of normal subjects (82,138). At the level of the esophageal hiatus, the fibers of the right crus extend almost directly anteriorly, and the most anterior portion may have a bulbous or nodular appearance that mimics left gastric adenopathy. (Fig. 10-14). The smaller left crus remains apposed to the aorta at this level. The posterior aspects of both crura lie against the pleural spaces.
to form the median arcuate ligament, surrounding the aorta immediately cephalic to the celiac trunk (110). This ligament can be imaged as a thin soft tissue stripe crossing the aorta; in some cases, it can produce deformity of the celiac trunk (Fig. 10-13) (134). The right crus is larger and originates lower than the left; it extends to the left of midline. As the crura continue upward, the right crus divides to enclose the esophagus within the esophageal hiatus. As the esophagus enters the stomach, both structures are relatively firmly tethered to the fissure for the ligamantum venosum
by the gastrohepatic ligament. As a result, a portion of the stomach wall runs in a transverse plane, and therefore appears thicker than the rest of the stomach on transverse sections (28). This “pseudomass” has been noted in 25% to 30% of normal subjects (82,138). At the level of the esophageal hiatus, the fibers of the right crus extend almost directly anteriorly, and the most anterior portion may have a bulbous or nodular appearance that mimics left gastric adenopathy. (Fig. 10-14). The smaller left crus remains apposed to the aorta at this level. The posterior aspects of both crura lie against the pleural spaces.
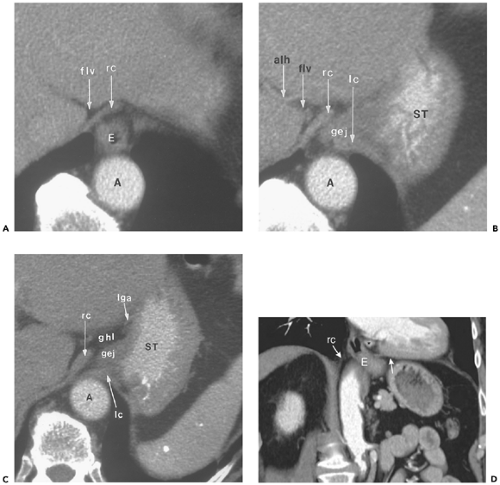 Figure 10-9 The esophageal hiatus. A: Section obtained immediately above the gastroesophageal junction shows the right diaphragmatic crus (rc) adjacent to the fat in the fissure for the ligamentum venosum (flv). The esophagus (E) passes between the aorta (A) and right crus as it courses from middle mediastinum into the abdomen. B: Section obtained 7 mm caudal to (A) shows the abdominal segment of the esophagus (gej) as it joins the stomach (ST) between the right (rc) and left (lc) crura. Of incidental note is an accessory left hepatic artery (alh), arising as a branch of the left gastric artery and coursing through the fissure for the ligamentum venosum (flv). C: Section obtained 7 mm caudal to (B) shows approximation of the right crus (rc) to the left (lc), effectively closing the esophageal hiatus. The left crus of the diaphragm remains in contact with the anterior surface of the aorta (A). Note that there has been an increase in the volume of fat within the gastrohepatic ligament (ghl) adjacent to the lesser curvature of the stomach (ST). lga, Left gastric artery; gej, gastroesophageal junction. D: Coronal reformatted image in another patient depicts the esophagus (E) coursing obliquely from the thorax into the abdominal cavity. Fibers of the right diaphragmatic crus (rc) sweep to the left to enclose the esophagus at the hiatus. Arrow, left leaf of the diaphragm; asterisk, paraesophageal lymph node. |
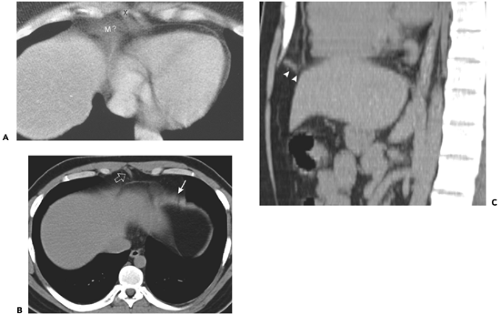 Figure 10-10 Appearance of anterior leaflets of the diaphragm. A: Pseudomass caused by anterior diaphragm. In this patient, the central tendon of the diaphragm is at a level close to the xiphoid process (X). In this setting, the broad muscles of the anterior diaphragm course in the same plane as the scan section, producing a pseudomass (M?) adjacent to the pericardium. B: In this individual, a portion of the anterior left hemidiaphragm (arrow) is imaged tangentially, simulating a mass. The fibers comprising the sternal origin of the diaphragm (open arrow) create a triangular soft tissue density. C: Reformatted sagittal image near the midline shows the fibers originating from the sternum, and extending posteriorly to join the central tendon (arrowheads). |
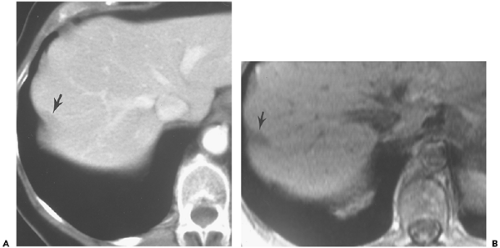 Figure 10-11 Diaphragmatic muscle mimicking liver lesion. A: Computed tomography section obtained at a level inferior to the dome of the diaphragm shows a peripheral band-like low-attenuation defect (arrow). B: Magnetic resonance imaging section in the same location shows a low-intensity structure (arrow) at the liver periphery. Serial images demonstrated its association with a rib. |
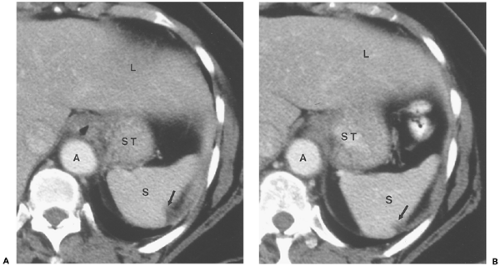 Figure 10-12 Splenic indentation from diaphragmatic slip. A: Section near the gastroesophageal junction shows a fat-containing notch (arrow) on the posterolateral aspect of the spleen (S). A, Aorta; ST, stomach; L, liver. B: Section obtained 5 mm inferior to (A) shows the soft tissue attenuation diaphragmatic slip passing through the splenic notch as it becomes continuous with the diaphragm. S, Spleen; A, aorta; ST, stomach; L, liver. |
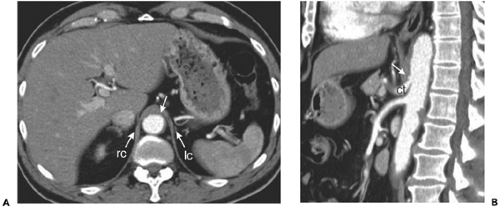 Figure 10-13 Median arcuate ligament. A: Axial section obtained just above the celiac trunk shows the median arcuate ligament (arrow) immediately anterior to the aorta, extending between the left (lc) and right crus (rc). B: Reformatted sagittal image obtained at end expiration shows the proximity of the celiac trunk (ct) with the median arcuate ligament (arrow). In some patients, end-expiratory imaging produces apparent occlusion of the proximal celiac artery. |
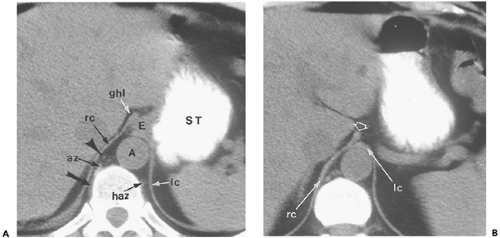 Figure 10-14 Retrocrural space. A: Section obtained at the gastroesophageal junction shows the abdominal segment of the esophagus (E) passing obliquely through the hiatus between the right (rc) and left crura (lc). The retrocrural space defined by the crural fibers is a continuation of the mediastinum. It contains the aorta (A); the azygous (az) vein, which on this section receives an intercostal vein (arrow); the hemiazygous (haz) vein; a variable amount of fat; the thoracic duct and lymph nodes (arrowhead); and part of the sympathetic trunk. At this level and the next, there is continuity between the retrocrural space and the abdominal contents, namely, the esophagus and gastrohepatic ligament (ghl). B: Section obtained 16 mm below (A). The left (lc) and right crura (rc) have reapposed below the esophageal hiatus. A small bulbous projection from the right crus (open arrow) projects into the region of the gastrohepatic ligament; this can mimic a node if its continuity with the remainder of the crus is not appreciated. |
The two crura enclose a retrocrural space anterior to the upper lumbar vertebral bodies (127). The aorta is the major component of this space, but the thoracic duct and azygos/hemiazygos veins also lie within it. The cisterna chyli is very frequently depicted as a bulbous structure of near water attenuation on the right side of the retrocrural space (Fig. 10-15) (46,112). The retrocrural space connects the posterior mediastinum with the abdominal retroperitoneum; pathologic processes in this space are confined on cephalic sections near the diaphragm, but below the median arcuate ligament, may escape anteriorly into the space anterior to the great vessels. Caudal to the median arcuate ligament, the vertebral origins of the diaphragm are seen as fusiform soft tissue densities lying on the lateral surface of the vertebral body anterior to the psoas muscles. On more caudal sections, the crura (most commonly the larger right crus) can be very nodular in appearance, so that they mimic paraaortic lymph nodes (155).
The medial and lateral arcuate ligaments course over the psoas and quadratus lumborum muscles to fuse with the diaphragm (99,109). The medial arcuate ligament extends from the lateral margin of the lumbar spine (L1) vertebral body to the transverse process of L1. The lateral arcuate ligament originates from the transverse process of the L1 vertebral body and inserts on the twelfth rib. In about 5% of normal subjects, the lateral arcuate ligament can appear very nodular (131), and knowledge of its appearance is helpful to avoid confusing it with pathology (Fig. 10-16).
Hernias tend to occur at specific places in the diaphragmatic surface. Anterior (Morgagni) hernias are retrosternal or positioned just lateral to the xiphoid on either side (34); their location may be the result of weakness at the site of penetration of the diaphragm by the superior epigastric vessels. Posterior (Bochdalek) hernias represent incomplete closure of the pleuroperitoneal canal (41). They are depicted on cross-sectional imaging with higher than expected frequency and are at least as likely on the right as
on the left (97) Both can contain abdominal fat or portions of subdiaphragmatic viscera.
on the left (97) Both can contain abdominal fat or portions of subdiaphragmatic viscera.
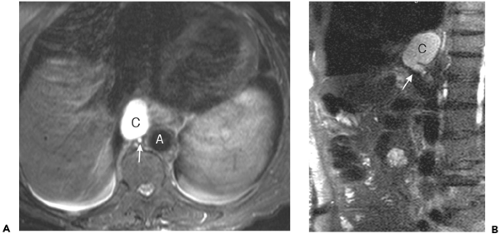 Figure 10-15 Cisterna chyli. A: T2-weighted magnetic resonance axial image through the thoracolumbar junction shows a large fluid-filled structure (C) just to the right of the aorta (A) and immediately anterior to the hemiazygos vein (arrow). B: Sagittal image in the same subject shows the junction of the dilated right lumbar trunk (arrowhead) with the cisterna chyli (C). |
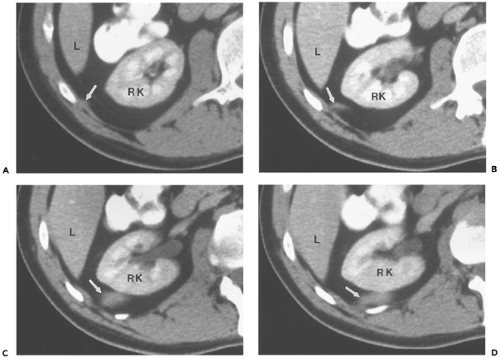 Figure 10-16 Lateral arcuate ligament. A–D: Serial sections beginning just inferior to the renal hilus show the lateral arcuate ligament (arrow) extending from a lateral position near its attachment to the rib (in [A]) to a more posterior position immediately behind the right kidney (RK) (in [D]). On the inferior segments, the ligament is thicker and broader than at its attachment, and mimics a mass. L, Liver. |
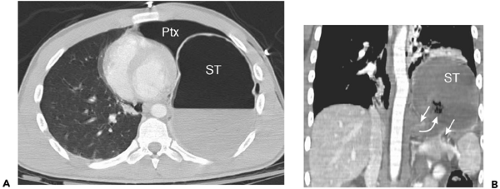 Figure 10-17 Traumatic rupture of the diaphragm. A: Axial section through the lower thorax in a patient involved in a motor vehicle collision shows a large left pneumothorax (Ptx). The fundus of the stomach (ST) lies occupies the posterior hemithorax at this level. B: Reformatted coronal image in the same patient shows large gap (between arrows) in the left hemidiaphragm, with protrusion of the gastric fundus (curved arrow) into the thorax through the defect. ST, stomach. |
Cross-sectional imaging has proved useful in evaluating patients with traumatic injuries to the diaphragm (16,17,76,98,100,124). In vehicular trauma, the tears tend to cluster about the junction between the central tendon and the muscular portion of the diaphragm and are much more common on the left. The diagnosis is important, chiefly because of the morbidity of delayed visceral herniation with strangulation; multidetector CT (MDCT) with sagittal and coronal reconstruction has positive and negative predictive values of about 80% (Fig. 10-17).
Intraperitoneal Organs
Embryology
The logic underlying the anatomic distribution of normal intraabdominal viscera and the distribution of pathologic processes arising from them is related to their embryology. Specifically, an understanding of the embryology of the mesenteries and the growth and development of intramesenteric organs provides a basis for understanding the complex nature of the peritoneal spaces in the upper abdomen, the formation of the abdominal ligaments (which play a critical role in defining the course of pathologic processes), and the genesis of the anterior pararenal space. Moreover, the distribution and patterns of extension of fluid collections in the retroperitoneum are explained by the same embryologic processes.
Embryologically, the gut develops from the yolk sac; it is suspended from the anterior and posterior body walls by ventral and dorsal mesenteries, which form the lateral boundaries of the developing alimentary tube. Early in fetal life, important organs develop in the mesenteries of the caudal part of the foregut. The dorsal mesogastrium is the site of the developing spleen and dorsal pancreas, whereas the liver expands within the ventral mesogastrium, along with the ventral pancreas (Fig. 10-18). Major fetal arteries course anteriorly through the dorsal mesenteries from the aorta to supply the developing gut and its newly developing organs. As the liver grows, the ventral mesogastrium thins greatly; the anterior part of it, which in the adult contains the obliterated umbilical vein (ligamentum teres), becomes the falciform ligament. In the adult, this structure is depicted as a midline triangular fat collection containing a fibrous cord that is the remnant of the obliterated umbilical vein. It enters the liver substance at the fissure for the ligamentum teres, forming an incomplete separation between the medial and lateral segments of the left hepatic lobe.
The dorsal part of the ventral mesogastrium (between the liver and the gut) also thins appreciably to form the lesser omentum. In the adult, this structure, with its accompanying fat, stretches between the lesser curvature of the stomach and the fissure for the ligamentum venosum, incompletely separating the caudate lobe from the left lateral segment. Within this gastrohepatic ligament are the left gastric artery and coronary vein, and part of the celiac lymph node chain (6,12) (Fig. 10-19). In some patients, a portion of the caudate lobe (the papillary process) extends between the lesser curve of the stomach and the portal vein and mimics lymphadenopathy (8).
More caudally, the ventral mesoduodenum forms the rest of the lesser omentum, the hepatoduodenal ligament (153,159). This structure encloses the portal vein, hepatic artery, common hepatic/common bile duct, and the hepatic group of celiac lymph nodes.
The spleen grows in the leaves of the dorsal mesogastrium, just posterior to the stomach. The ventral part of this
mesentery, the gastrosplenic ligament, ultimately becomes part or all of three very important upper abdominal structures: the greater omentum, the transverse mesocolon, and the gastrosplenic ligament (Fig. 10-20). On sections near the spleen, the ligament is short and is situated posteriorly; it transmits the short gastric vessels behind the inferior recess of the lesser sac (Fig. 10-21). Inferiorly, the peritoneum inside the lesser sac greatly elongates the dorsal mesentery. The apposed redundant surfaces fuse to form the gastrocolic ligament, or greater omentum, which can be recognized by the presence of the gastroepiploic vessels, which arise near the hilus of the spleen (Fig. 10-22) (24). Dilation of the gastroepiploic veins is a very common sign of splenic vein occlusion, frequently a consequence of pancreatitis. A small posterior part of the redundant dorsal mesentery fuses with the dorsal mesentery of the transverse colon to create the multi-layered transverse mesocolon. The root of this structure lies posterior and inferior to the lesser sac and can also be identified by its vessels: the middle colic artery and vein (21) (Fig. 10-23).
mesentery, the gastrosplenic ligament, ultimately becomes part or all of three very important upper abdominal structures: the greater omentum, the transverse mesocolon, and the gastrosplenic ligament (Fig. 10-20). On sections near the spleen, the ligament is short and is situated posteriorly; it transmits the short gastric vessels behind the inferior recess of the lesser sac (Fig. 10-21). Inferiorly, the peritoneum inside the lesser sac greatly elongates the dorsal mesentery. The apposed redundant surfaces fuse to form the gastrocolic ligament, or greater omentum, which can be recognized by the presence of the gastroepiploic vessels, which arise near the hilus of the spleen (Fig. 10-22) (24). Dilation of the gastroepiploic veins is a very common sign of splenic vein occlusion, frequently a consequence of pancreatitis. A small posterior part of the redundant dorsal mesentery fuses with the dorsal mesentery of the transverse colon to create the multi-layered transverse mesocolon. The root of this structure lies posterior and inferior to the lesser sac and can also be identified by its vessels: the middle colic artery and vein (21) (Fig. 10-23).
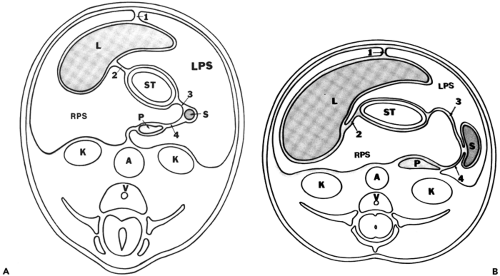 Figure 10-18 Mesenteries attached to the stomach and the developing intramesenteric viscera. Adapted from reference 74. A: Schematic drawing of a section obtained in an embryo, near the end of the fifth week of development. The stomach (ST) is supported by two major mesenteries, ventral and dorsal. Developing within the ventral mesentery, and distorting its surface, the liver (L) grows chiefly into the right peritoneal space (RPS). Maternal blood courses through the ventral part of the ventral mesentery, which becomes the falciform ligament (1). The dorsal portion of this ventral mesentery (2) contains the left gastric artery and coronary vein and, more caudally, the hepatic artery, portal vein, and biliary duct within its leaves. This mesentery will become the lesser omentum. The spleen (S) takes shape in the ventral part of the dorsal mesentery; the gastrosplenic ligament (3) formed from it carries the short gastric vessels. Although the head of the pancreas (P) arises in the dorsal mesoduodenum, its tail grows in a cephalic direction to occupy the dorsal mesogastrium within the splenorenal ligament (4); (A) aorta; (K) kidney; (V) vertebral body; (LPS) left peritoneal space. B: Approximately 1 week later, the rapid hepatic growth forces considerable rotation of the stomach (ST) and attached lesser omentum (2). Meanwhile, the pancreatic tail (P) has fused to the dorsal body wall, reducing the posteromedial extent of the left peritoneal cavity (LPS). This line of fusion generally continues along the splenorenal ligament to form a posteromedial splenic “bare” or nonperitonealized area. In some patients, this fusion does not occur, and peritoneum extends behind the posterior pancreatic tail. In this condition, the spleen is on a mesentery of variable length and can “wander” within the peritoneal cavity; 1, falciform ligament; 3, gastrosplenic ligament; 4, splenorenal ligament; A, aorta; K, kidney; V, vertebral body; L, liver; S, spleen. |
The dorsal segment of the dorsal mesogastrium extends posteriorly from the spleen to enclose the pancreatic tail and splenic vessels, and then fuses with the posterior body wall, joining with the fascia anterior to the left kidney. This
forms the splenorenal ligament (failure of this fusion process leads to the rare clinical condition of “wandering spleen”) (3). Because of this same fusion, part of the posteroinferior surface of the spleen (adjacent to the upper pole of the left kidney) is not peritonealized. The splenic bare area limits the distribution of fluid in the perisplenic space (144) (Fig. 10-24).
forms the splenorenal ligament (failure of this fusion process leads to the rare clinical condition of “wandering spleen”) (3). Because of this same fusion, part of the posteroinferior surface of the spleen (adjacent to the upper pole of the left kidney) is not peritonealized. The splenic bare area limits the distribution of fluid in the perisplenic space (144) (Fig. 10-24).
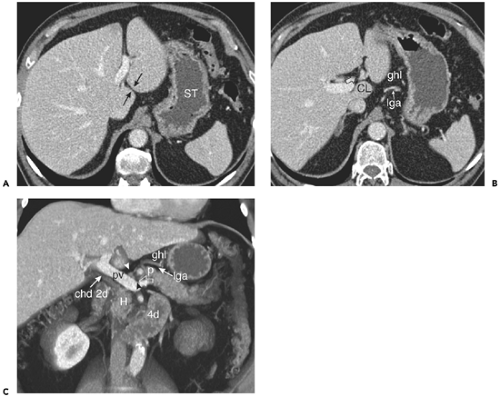 Figure 10-19 The gastrohepatic and hepatoduodenal ligaments. A: The gastrohepatic ligament extends between the lesser curvature of the stomach (ST) and the fissure for the ligamantum venosum (between arrows). It contains branches of the left gastric artery and coronary veins. B: Axial section obtained 20 mm inferior to (A) shows the course of the left gastric artery (lga) through the fat-containing gastrohepatic ligament (ghl). Just anterior to the caudate lobe (CL) is a slightly enlarged lymph node (open arrow). C: Reformatted volume rendered image shows the structures of the lesser omentum. The lower portion (hepatoduodenal ligament) contains the portal vein (pv), common hepatic duct (chd) and hepatic artery (black arrowhead). A small hepatic chain lymph node is present (white arrowhead). To the left and superiorly, the gastrohepatic ligament (ghl) contains the left gastric artery (lga). Note drainage (open arrow) of the coronary vein into the portal vein near the splenoportal confluence. A portion (called the tuber omentale) of the body of the pancreas (p) protrudes into the gastrohepatic ligament. 2d, Descending duodenum; 4d, fourth portion of the duodenum; H, pancreatic head. |
The ventral mesentery of the small intestine and colon distal to the midduodenum resorbs and allows free communication of the right with the left peritoneal spaces. The dorsal mesentery of the small intestine simply grows to keep pace with the rapid growth of the small bowel it supports. The dorsal mesentery of the colon, however, has a role to play in forming both the upper peritoneal spaces and the retroperitoneal anterior pararenal space (Fig. 10-25). Initially, the colon is a straight tube lying parallel to the long axis of the fetus and supplied by the superior mesenteric artery via its dorsal mesentery. With time, the colon elongates and eventually, with the small intestine, herniates through the umbilical defect into the extraembryonic coelomic cavity.
When the colon returns to the abdominal cavity, it undergoes rotation (74,94). Its distal part, which will become the descending colon in the adult, maintains the same orientation parallel to the long axis of the fetus. The midportion of the colon rotates 90 degrees (in a counterclockwise direction, as viewed looking down on the fetus) to lie perpendicular to the long axis. The proximal colon, which will become the cecum and ascending portions, undergoes yet another 90-degree rotation in the same counterclockwise mode, so that the original left side of its dorsal mesentery now points to the right side of the abdomen. Finally, the descending colon and its dorsal mesentery rotate (in a clockwise direction, as viewed looking from fetal foot to fetal head), so that its left side fuses to the anterior fascial lining
of the left kidney. In a similar fashion, the ascending colon and its mesentery rotate in a counterclockwise mode, fusing to the anterior fascial lining of the right kidney. The transverse colon does not fuse to the body wall; instead, it “flops” inferiorly, so that the left side of its dorsal mesentery faces anteriorly. It is this surface that fuses to the posterior surface of the gastrosplenic ligament (described previously) to form the transverse mesocolon. Note that as a result of these rotations, the left surface of the dorsal colonic mesentery fuses with the left anterior renal fascia, the gastrosplenic mesentery, and the right anterior renal fascia. This forms an intercommunicating system of planes through which pathologic fluid collections (notably pancreatitis) can be distributed throughout the abdomen (2,48).
of the left kidney. In a similar fashion, the ascending colon and its mesentery rotate in a counterclockwise mode, fusing to the anterior fascial lining of the right kidney. The transverse colon does not fuse to the body wall; instead, it “flops” inferiorly, so that the left side of its dorsal mesentery faces anteriorly. It is this surface that fuses to the posterior surface of the gastrosplenic ligament (described previously) to form the transverse mesocolon. Note that as a result of these rotations, the left surface of the dorsal colonic mesentery fuses with the left anterior renal fascia, the gastrosplenic mesentery, and the right anterior renal fascia. This forms an intercommunicating system of planes through which pathologic fluid collections (notably pancreatitis) can be distributed throughout the abdomen (2,48).
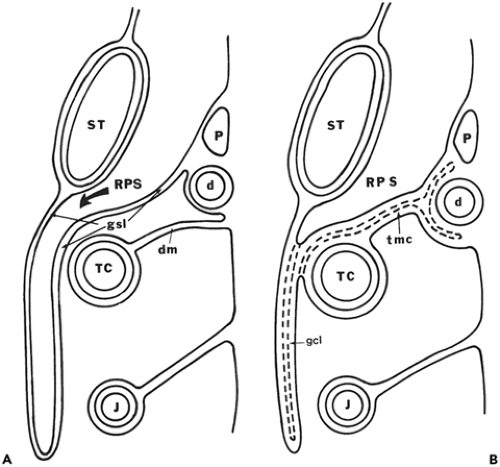 Figure 10-20 Formation of the gastrocolic ligament. Adapted from reference 74. A: Schematic drawing of a sagittal section through a developing embryo. Growth of the right peritoneal space (RPS) behind the stomach (ST) has greatly elongated the gastrosplenic ligament (GSL), so that it hangs like a drape over the transverse colon (TC) and its attached dorsal mesentery (dm); P, pancreas; d, duodenum; J, jejunum. B: Fusion occurs between anterior and posterior surfaces of the gastrosplenic ligament, obliterating part of the right peritoneal space (RPS) and forming the gastrocolic ligament (gcl). The posterior surface of the fused ligament in turn fuses to the transverse colon (TC) and its dorsal mesentery to form the adult transverse mesocolon (tmc); P, pancreas; d, duodenum; J, jejunum; ST, stomach. |
Peritoneal Spaces
Complex rotation and fusion of mesenteric structures alter the peritoneal anatomy; growth of intramesenteric organs, especially the liver, further distorts its appearance. A schematic overview of the distribution of peritoneal spaces in the adult is shown in Fig. 10-26.
The left peritoneal space in the adult can be divided into four compartments (all of which intercommunicate): two perihepatic spaces, anterior and posterior; and two subphrenic spaces, anterior and posterior (Fig. 10-27). The left anterior perihepatic space is confined on the right by the falciform ligament. The left posterior perihepatic space follows the undersurface of the lateral segment of the left hepatic lobe, and extends deep into the fissure for the ligamentum venosum (7,91,122,147). The left anterior subphrenic space is the continuation of the anterior perihepatic space (53). It courses over the gastric fundus just posterior to the anterior leaf of the left hemidiaphragm. Large collections in the left peritoneal space can extend superiorly through the diaphragmatic hiatus into the mediastinum (45), or posterolaterally into the posterior subphrenic (perisplenic) space. This space covers the superior and inferolateral surfaces of the spleen,
but is confined medially by the splenorenal ligament and the bare area of the spleen. Below the spleen, the phrenicocolic ligament separates the perisplenic space from the rest of the peritoneal cavity (Fig. 10-28).
but is confined medially by the splenorenal ligament and the bare area of the spleen. Below the spleen, the phrenicocolic ligament separates the perisplenic space from the rest of the peritoneal cavity (Fig. 10-28).
The right peritoneal space has two major divisions: the perihepatic space and the lesser sac (omental bursa). The right perihepatic space occupies the broad expanse between the anterior and lateral surfaces of the right lobe of the liver and the right hemidiaphragm. It is confined on the left and anteriorly by the falciform ligament, and posteriorly by the right coronary ligament, which marks the lateral aspect of the bare area of the liver (120) (Fig. 10-29). On successively caudal scans, the laterally placed peritoneal recess extends more and more medial as the bare area becomes smaller; at the level of the upper right renal pole, it turns anterior to occupy a space between the posterior liver surface and the anterior renal fascia (121). This posterior recess is named the hepatorenal fossa or the Morison pouch. On scans below the gallbladder fossa, the perihepatic space completely encircles the right hepatic segments (Fig. 10-30).
The anatomy of the lesser sac attests to its complex embryologic origin. As described previously, it is a part of the right peritoneal space that is displaced as the liver grows to fill the right upper abdomen. The right peritoneal space then moves medially behind the gastrohepatic ligament and stomach and is enfolded by the redundant gastro-splenic ligament. As a result, collections in the lesser sac are difficult to access by percutaneous puncture; that space is completely surrounded by mesenteries and intramesenteric organs (96).
In the adult, two major recesses are present. The superior recess completely encloses the medial surface of the caudate lobe (117) (Fig. 10-31). At the porta hepatis, the superior recess lies just posterior to the portal vein; on more cephalic sections, it lies immediately behind the lesser omentum deep within the fissure for the ligamentum venosum
and follows the caudate lobe surface posteriorly and to the right, extending almost to the inferior vena cava. Near the diaphragm, the posterior part of this space lies adjacent to the right diaphragmatic crus, just inferior to the abdominal segment of the esophagus (4). The inferior recess of the lesser sac (see Fig. 10-29) lies between the stomach and the visceral surface of the spleen; on lower sections, it separates the stomach from the pancreas and the transverse mesocolon (30,66). Part of this space may potentially extend between the leaves of the greater omentum (Fig. 10-32).
and follows the caudate lobe surface posteriorly and to the right, extending almost to the inferior vena cava. Near the diaphragm, the posterior part of this space lies adjacent to the right diaphragmatic crus, just inferior to the abdominal segment of the esophagus (4). The inferior recess of the lesser sac (see Fig. 10-29) lies between the stomach and the visceral surface of the spleen; on lower sections, it separates the stomach from the pancreas and the transverse mesocolon (30,66). Part of this space may potentially extend between the leaves of the greater omentum (Fig. 10-32).
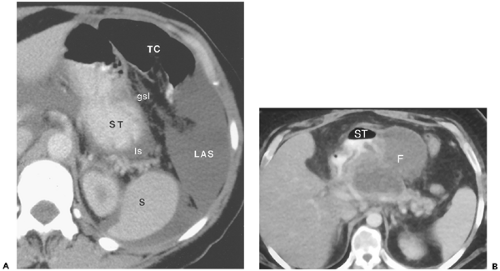 Figure 10-21 The gastrosplenic ligament. A: Computed tomography section through the upper abdomen in a patent with ascites shows fluid-filled peritoneal spaces outlining the gastrosplenic ligament (gsl). The gastrosplenic ligament is the fat-containing structure between the left anterior subphrenic space (LAS) and the inferior recess of the lesser sac (ls). The serpentine structures within this fat are the short gastric arteries, which arise from the splenic artery and supply the greater curvature of the stomach (ST). Embryologically, the gastrosplenic ligament forms part of the transverse mesocolon and all of the greater omentum. S, spleen. B: Fluid in the gastrosplenic ligament. In this patient with pancreatitis, a large fluid collection (F) within the gastrosplenic ligament distorts the posterolateral wall of the stomach (ST). |
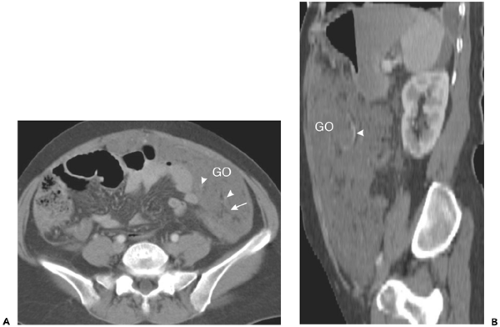 Figure 10-22 The greater omentum. A: Axial section in a patient with widespread peritoneal metastases shows extensive soft tissue involvement of the greater omentum (GO). Omental fat (arrow) and gastroepiploic vessels (arrowhead) persist. B: Sagittal image in the same patient demonstrates the cephalocaudal extent of the omentum (GO) and the gastroepiploic vessels (arrowhead) coursing within the small amount of preserved omental fat. |
Posterior Recesses of the Peritoneum
The precise distribution of the most posterior recesses of the peritoneal spaces depends on the completeness of fusion between the dorsal mesenteries and the posterior abdominal retroperitoneum. When fusion is incomplete, peritoneum may extend posterior to the renal margin on either side. Because mobile structures in the peritoneal cavity can inhabit any portion of the peritoneal space, it is possible for the colon or spleen to lie in a retrorenal or retrogastric position (59,105). Clearly, this variation may be a considerable hazard to patients undergoing percutaneous renal procedures.
Extraperitoneal Organs, Spaces, and Planes
The extraperitoneal spaces include all structures that lie between the external part of the transversalis fascia and the peritoneal lining. There are two major components to the extraperitoneal spaces: the abdominal retroperitoneum, and the pelvic perivesical space. The latter will be considered in the portion of this chapter that deals with pelvic anatomy.
There is really no consistent definition of the region referred to as the abdominal retroperitoneum. The most commonly accepted definition refers to the retroperitoneum as the fat-filled compartment that lies between the parietal peritoneum and the transversalis fascia. However, imaging observations suggest that the retroperitoneum is not a totally separate compartment, but is continuous with portions of the anterior abdominal wall, with the pelvic perivesical extraperitoneal space (27,43), and with the mesenteries discussed in the section on peritoneum.
The purpose of this section is to incorporate these clinical observations of the distribution of retroperitoneal fluid with the classically described anatomy. Rapidly evolving fluid collections (for example, those seen in severe pancreatitis) are too large to be maintained in their space of origin (29,89). Surprisingly, even aggressive processes take very well-defined routes to escape: they either dissect into an adjacent mesentery (a process known as subperitoneal spread) (106,123) or they evacuate into fascial planes that lie between the retroperitoneal spaces (intrafascial spread). It is the latter that explains the connections observed between the diaphragm superiorly and the extravesical pelvic spaces inferiorly (2,15,48,69,93,103,142,158).
Great Vessel Space
The aorta and its major visceral branches as well as the inferior vena cava and its tributaries course within an ill-defined space anterior to the vertebral bodies (Fig. 10-33). This space is a caudal continuation of the posterior mediastinum, but no discrete fascial planes confine it, and processes may extend from it to involve other retroperitoneal spaces. Paraaortic and paracaval lymph nodes accompany the entire course of
the abdominal aorta and vena cava. The diaphragmatic crura enclose the upper part of the abdominal aorta, ending in a fibrous arch, the median arcuate ligament, immediately cephalic to the level of the celiac trunk. The origin of the superior mesenteric artery is 1 to 2 cm caudal; the course of this artery parallels the aorta over several centimeters. Approximately 1 to 2 cm more caudal, the renal arteries exit the aorta laterally or posterolaterally to enter the renal hila. The right renal artery passes behind the inferior vena cava (IVC) in its course. Above the aortic bifurcation 1 to 2 cm, the inferior mesenteric artery exits the left anterolateral surface of the aorta and gives off left colic, sigmoid, and superior rectal branches.
the abdominal aorta and vena cava. The diaphragmatic crura enclose the upper part of the abdominal aorta, ending in a fibrous arch, the median arcuate ligament, immediately cephalic to the level of the celiac trunk. The origin of the superior mesenteric artery is 1 to 2 cm caudal; the course of this artery parallels the aorta over several centimeters. Approximately 1 to 2 cm more caudal, the renal arteries exit the aorta laterally or posterolaterally to enter the renal hila. The right renal artery passes behind the inferior vena cava (IVC) in its course. Above the aortic bifurcation 1 to 2 cm, the inferior mesenteric artery exits the left anterolateral surface of the aorta and gives off left colic, sigmoid, and superior rectal branches.
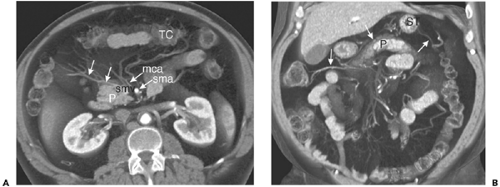 Figure 10-23 The transverse mesocolon. A: Volumetric axial reformatted image through the midabdomen shows the middle colic artery (mca) arising from the superior mesenteric artery (sma) and branching within the transverse mesocolon to supply the transverse colon (TC). Tributaries of the middle colic vein (arrows) here drain directly into the superior mesenteric vein (smv). P, pancreatic head and uncinate process. B: Coronal reformatted image in the same patient shows the transverse mesocolon spanning the entire upper abdomen. The middle colic veins (arrows) mark the position of the mesocolon, which lies immediately superior to the pancreas (P) and inferior to the stomach (ST).
Related posts:Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
 Get Clinical Tree app for offline access
Get Clinical Tree app for offline access

|