CHAPTER 13 Ventricles and Intracranial Subarachnoid Spaces
The brain lies within the skull submerged in a bath of cerebrospinal fluid (CSF). In the adult, the total volume of CSF is approximately 150 mL, of which about 125 mL is intracranial and 25 mL is intraspinal.1 Within the brain, the CSF fills four hollow spaces designated the left and right lateral ventricles, the third ventricle, and the fourth ventricle. The two lateral ventricles lie off the midline, within the left and right cerebral hemispheres. The third ventricle lies in the midline between the two thalami. The fourth ventricle lies immediately posterior to the brain stem at the center of the cerebellum (Figs. 13-1 to 13-3). In adults, the mean volume of the two lateral ventricles varies from 9 to 27.6 mL and is slightly larger in men than women.2–5 The mean volumes of the left and right lateral ventricles are estimated individually at 14.4 mL and 13.2 mL, respectively.3 The combined volume of the third ventricle, aqueduct, and fourth ventricle is estimated at 1.0 to 1.5 mL.4 Two additional compartments of CSF situated between the leaves of the septum pellucidum (cavum septi pellucidi) and between the fornices (cavum vergae) are occasionally considered to be fifth and sixth ventricles, respectively.
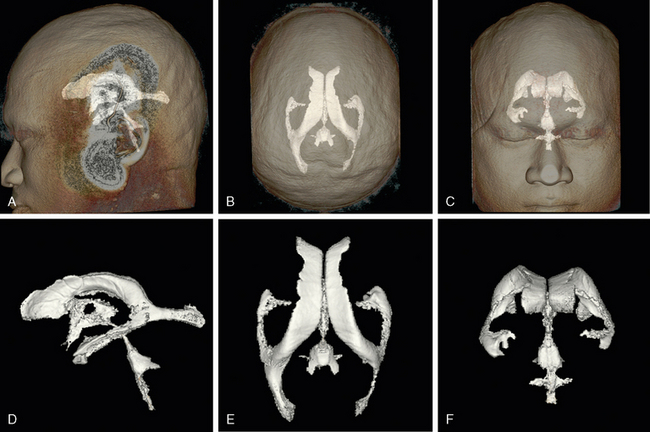
FIGURE 13-1 Ventricular system. Three-dimensional (3D) computer reformatted volumetric T2W MR images display the ventricles in lateral (A, D), superior (B, E), and anterior (C, F) views. A to C, Orientation of the ventricular system within the head, seen through the partially transparent skin surface. D to F, Magnified display of the isolated ventricles in the same orientation. See also Videos 13-1 and 13-2 at www.expertconsult.com for an interactive rotation.
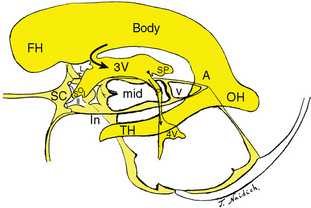
FIGURE 13-2 Schematic representation of the ventricular system in relation to the suprasellar cistern (SC), optic chiasm (II), incisura (In), midbrain (mid), vermis (v), and subarachnoid cisterns of the posterior fossa. Semilateral view. The lateral ventricle displays the frontal horn (FH), body (Body), atrium (A), occipital horn (OH), and temporal horn (TH). The lateral ventricle leads to the third ventricle through the foramen of Monro (curved black arrow). The anterior third ventricle (3V) displays the supraoptic (SO) and infundibular (I) recesses enclosing the optic chiasm (II). The posterior third ventricle displays a large suprapineal recess (SP) and small pineal recess (black arrowhead). The aqueduct (dotted black arrow) leads into the fourth ventricle (4V). Compare with Figure 13-1A and D.
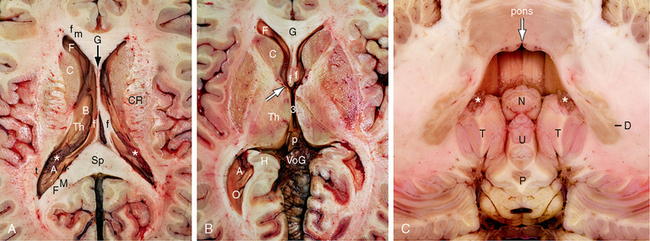
FIGURE 13-3 Ventricular system. Magnified axial plane gross anatomic specimens. A and B, Supratentorial ventricles. A, Slightly asymmetric axial section through the midlateral ventricles. At this level, the frontal horn (F) is delimited anteriorly and medially by the forceps minor (fm) that crosses through the genu (G) of the corpus callosum, delimited medially by the midline septum pellucidum (black arrow), laterally by the white matter surrounding the lateral angle of the ventricle, and inferolaterally by the caudate head (C). The body (B) of the lateral ventricle is delimited inferiorly by the superior surface of the thalamus (Th) laterally and by the fornix (f) medially. The atrium (A) is delimited anteriorly by the pulvinar of the thalamus (Th), superiorly and medially by the forceps major (FM) that crosses through the splenium (Sp) of the corpus callosum, and laterally by the tapetum (t). The choroid plexus (asterisk) emerges into the lateral ventricle at the lateral edge of the fornix. It forms the glomus of the choroid plexus in the atrium. CR, corona radiata. B, Axial section through the frontal horns (F), foramina of Monro (white arrow), and third ventricle (3). The anterior columns of the fornices (f) form the anteromedial walls of the foramina of Monro. The anterior nuclei of the thalami (Th) form the posterolateral walls of the foramina of Monro. The medial aspects of the thalami form the lateral walls of the third ventricle. The pineal gland (p) lies at the posterior end of the third ventricle and protrudes backward into the cistern of the vein of Galen (VoG). The occipital horns (O) extend posteriorly from the atria, very often asymmetrically. H, hippocampal formation. C, Infratentorial ventricle. In the axial plane, the fourth ventricle (white arrow) has the shape of a temple bell. The “top” of the bell is formed by the midline groove (also white arrow) in the floor of the fourth ventricle. To each side of midline, the upper curvatures are formed by the paired median eminences. The concave side walls of the fourth ventricle are formed by the superior and inferior cerebellar peduncles. The posterior wall is formed by the nodulus (N) of the vermis in the midline and the paired cerebellar tonsils (T) to each side. Because the midline fastigial recess of the fourth ventricle curves over the nodulus, it is seen in the midline superior to and medial to the paired posterior superior recesses that curve over the paramedial tonsils. The choroid plexus (white asterisks) of the fourth ventricle is a paired paramedian structure that lies atop the tonsil on each side and extends symmetrically from there into the foramen of Magendie and the paired foramina of Luschka. The dentate nuclei (D) lie to each side of the fourth ventricle. Also labeled are the uvula (U) and the pyramis (P) of the inferior vermis.
(See also the animations in Videos 13-1 and 13-2 at www.expertconsult.com.)
ANATOMY
Lateral Ventricles
The lateral ventricles are paired cavities lying to each side of the midline within the cerebral hemispheres (see Figs. 13-1 to 13-5). They are imperfectly symmetric in most cases and show definite asymmetry in 5% to 12% of neurologically intact patients.6 The asymmetry does not appear to correlate with handedness, age, gender, or function.2,6 As seen from the side, each lateral ventricle is roughly C shaped, open anteriorly. Each lateral ventricle shows prominent extensions away from the main body of the ventricle. These extensions are termed horns. The anterior extensions into the frontal lobes are the anterior (frontal) horns. The posterior extensions into the occipital lobes are the posterior (occipital) horns. The inferior extensions into the temporal lobes are the inferior (temporal) horns. Posteriorly on each side, the occipital horn and temporal horn unite with the posterior body to form a large, confluent CSF space designated the atrium (trigone) of the lateral ventricle.
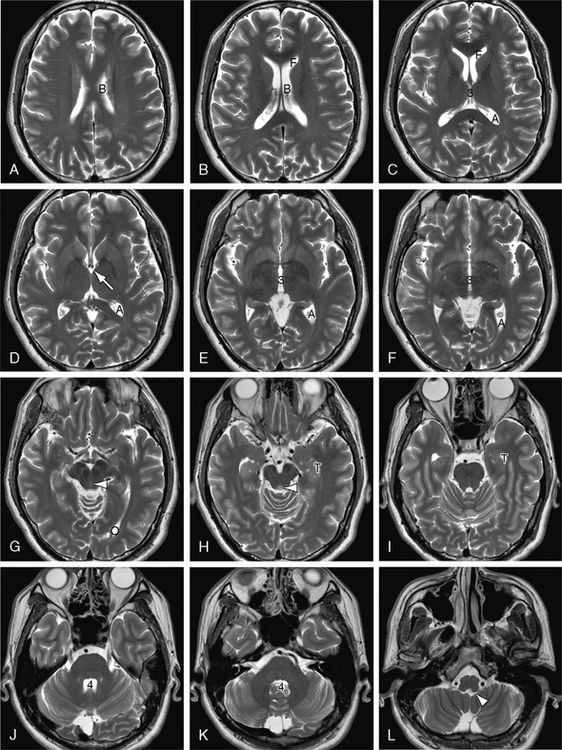
FIGURE 13-4 Ventricular system. A to L, Serial axial T2W MR images displayed from superior to inferior. In order from superior to inferior, serial axial sections display the body (B) of the lateral ventricle, then the frontal horns (F), atria (A), superior third ventricle (3), foramina of Monro (white solid arrow), mid third ventricle (3), occipital horn (O), aqueduct of Sylvius (white arrowhead in G and H), temporal horn (T), mid fourth ventricle (4), and foramina of Luschka (white arrowhead in L).
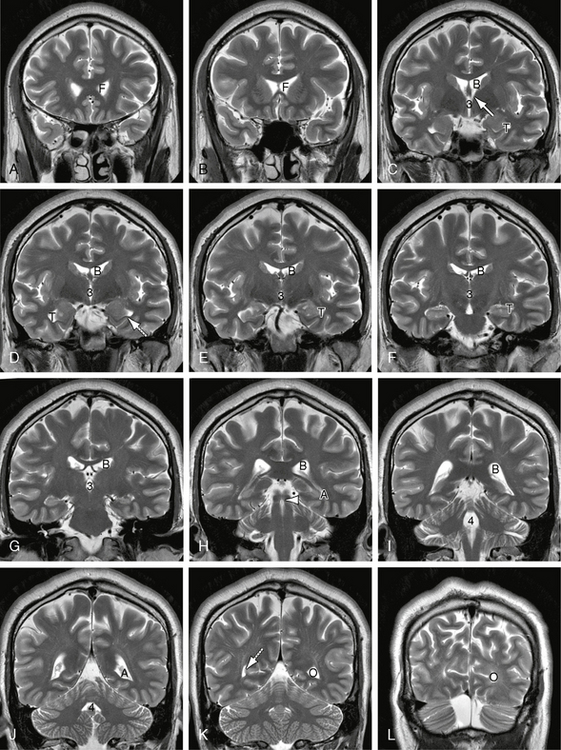
FIGURE 13-5 Ventricular system. A to L, Serial coronal T2W MR images displayed from anterior to posterior. In order from anterior to posterior, serial coronal sections display the frontal horns (F) of the lateral ventricles, the foramina of Monro (solid white arrow), the body (B) of the lateral ventricle, third ventricle (3), temporal horn (T), aqueduct of Sylvius (white arrowhead), fourth ventricle (4), atrium (A), and occipital horns (O). The collateral fissure of the temporal lobe buckles inward to indent the floor of the lateral ventricle, raising a prominence designated the collateral eminence (dotted white arrow in D). The calcarine sulcus and its junction with the parieto-occipital sulcus similarly indent the medial wall of the occipital horn, raising a medial bulge designated the calcar avis (dotted white arrow in K).
The occipital horns are the most asymmetric portion of the lateral ventricles and may be absent on one or both sides in normal individuals. They are delimited superiorly by the corpus callosum and the white matter fibers that cross through the splenium (forceps major). They are delimited laterally by the tapetum and medially by the subcortical white matter of the occipital lobes. The deep calcarine and parieto-occipital fissures on the medial surface of the occipital lobe often buckle the subcortical white matter laterally to indent the medial wall of the horn. This creates a variably prominent bulge on the medial surface of the occipital horn designated the calcar avis. When particularly large, the calcar avis may (nearly) amputate the posterior portion of the occipital horn. The occipital horns contain no intrinsic choroid plexus but may contain the dependent portion of a pedunculated glomus, unilaterally or bilaterally.
The temporal horns of the lateral ventricles extend inferolaterally into the medial temporal lobes. Their mean volume has been estimated to be 0.46 mL in normal subjects.3 The temporal horns are delimited inferomedially by the hippocampal formations, anterosuperiorly by the amygdalae, and laterally by the tapetum, optic radiations, and other white matter of the temporal lobes.7 The collateral sulcus on the inferior surface of the temporal lobe often buckles the subcortical white matter upward, into the floor of the temporal horn, to create a characteristic “collateral eminence” along the floor of the temporal horn (see Fig. 13-5) The anterior tips of the temporal horns have no intrinsic choroid plexus. The choroid plexus enters the temporal horns behind the temporal tip through the choroidal fissure along the superior medial wall of the temporal horn.
On each side, the choroid plexus of the body of the lateral ventricle is supplied by the lateral posterior choroidal arteries arising from the posterior cerebral artery. The choroid plexus of the temporal horns is supplied by the anterior choroidal arteries that arise from the supraclinoid internal carotid arteries. The anterior and posterior choroidal supplies meet near the atria of the ventricles and the glomera of the choroid plexus. The choroid plexus of the lateral ventricles is drained by the superior choroidal veins.
Foramina of Monro
The paired foramina of Monro interconnect the two lateral ventricles with the midline third ventricle (see Figs. 13-3 to 13-5; see also Chapter 11). The mean sagittal diameter of each foramen is 2.9 mm, and the mean vertical height is 5.1 mm.8 The anteromedial walls of the foramina are formed by the two anterior columns of the fornices. The posterolateral walls are formed by the anterior nuclei of the thalami. On each side, the choroid plexus of the lateral ventricle extends through the foramen of Monro to become continuous with the choroid plexus in the roof of the third ventricle.
Third Ventricle
The third ventricle is the midline cavity that connects with the lateral ventricles anterosuperiorly via the foramina of Monro and connects with the fourth ventricle posteroinferiorly via the cerebral aqueduct of Sylvius (Figs. 13-2 and 13-6 to 13-9; see also Chapter 11). It is ellipsoidal and 4 to 11 mm wide.9 The third ventricle is delimited anteriorly by the lamina terminalis and the anterior commissure; anterosuperiorly by the paired foramina of Monro; superiorly by the velum interpositum (velum transversum) and the habenulae; posteriorly by the pineal gland, the hippocampal commissure, and the posterior commissure; laterally by the thalamus and hypothalamus; and inferiorly by the optic chiasm and hypothalamus. The major hypothalamic structures related to the third ventricle include the infundibulum, the tuber cinereum, and the paired mammillary bodies. The striae medullares thalamorum course anteroposteriorly along the lateral border of the roof of the third ventricle, just where the roof inserts laterally into the medial walls of the thalami. The lamina terminalis is a thin neuroglial membrane that extends from the rostrum of the corpus callosum and anterior commissure superiorly down to the superior surface of the optic chiasm inferiorly. It is lined on the ventricular surface by ependyma and on the rostral surface by pia-arachnoid. Embryologically, the lamina terminalis is the “top” of the early neural tube and the site of the anterior neuropore. In most cases the cavity of the third ventricle is crossed by the massa intermedia (synonyms: interthalamic connection, interthalamic adhesion). The massa intermedia is composed of thalamic gray matter, not white matter, and varies widely in size from 1 × 2 mm to 1 × 2 cm.
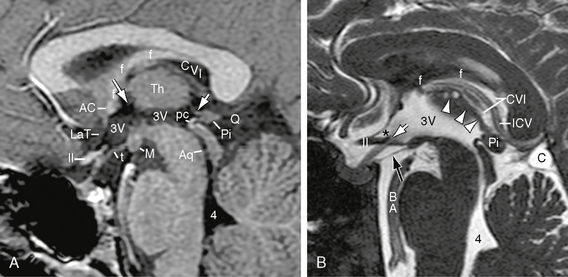
FIGURE 13-6 Third ventricle. A, Midsagittal contrast-enhanced T1W MR image. The walls of the third ventricle (3V) are formed anteriorly by the anterior commissure (AC) and lamina terminalis (LaT); anteroinferiorly by the optic chiasm (II) and anterior recesses of the third ventricle; inferiorly by the tuber cinereum (t), mammillary bodies (M), and midbrain; posteriorly by the posterior commissure (pc), pineal gland (Pi), and habenular commissure (short white arrow); laterally by the thalamus (Th); and superiorly by the membranous roof of the third ventricle. Normally, the curvature of the floor of the third ventricle continues smoothly into the aqueduct (Aq) and along the floor of the fourth ventricle (4). The foramen of Monro (long white arrow) lies between the anterior column of the fornix (f) and the anterior nucleus of the thalamus (Th) on each side. The cistern of the velum interpositum (CVI) extends forward from the quadrigeminal plate cistern (Q) almost to the foramen of Monro between the fornix (f) above and the roof of the third ventricle below. B, Midsagittal T2W FIESTA (fast imaging employing steady-state acquisition) MR image. The supraoptic recess (asterisk) lies anteriorly between the lamina terminalis and the upper surface of the chiasm (II). The infundibular recess (white arrow) lies farther posteriorly, behind the chiasm, and extends down into the infundibulum. The diencephalic leaf (black arrow) of the membrane of Liliequist extends from the dorsum sellae and posterior clinoid processes inferiorly to insert into the mammillary bodies superiorly (20% at the anterior margin of the mammillary bodies, 33% at their apex, and 47% at their posterior edge).21 This leaf separates the chiasmatic cistern above from the interpeduncular cistern below. Multiple linear defects within the interpeduncular fossa indicate the thalamoperforating branches of the basilar and proximal posterior cerebral arteries. Superiorly, the choroid plexus (arrowheads) of the third ventricle gives the roof an irregular contour and presents small choroid plexus cysts. Superior to that, the cistern of the velum interpositum (CVI) shows a well-defined upper margin along the fornix (f) and a well-defined lower margin along the roof of the third ventricle. It contains the internal cerebral vein (ICV). A small loculated arachnoid cyst (C) manifests very high signal intensity when compared with the adjacent subarachnoid fluid.
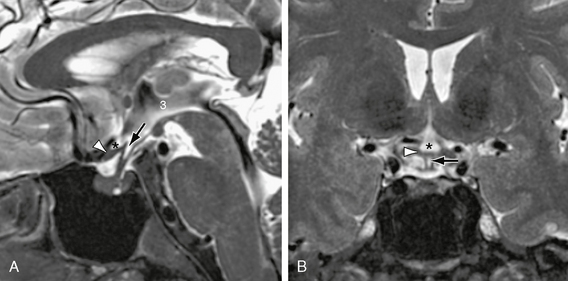
FIGURE 13-7 Anterior third ventricle. A, Midsagittal T2W MR image displays the supraoptic recess (asterisk) and the infundibular recesses (arrow) of the third ventricle (3) enclosing the optic chiasm (white arrowhead). B, In coronal section, the supraoptic recess (asterisk) resembles an Erlenmeyer flask resting on the chiasm. The infundibular recess (arrow) resembles a pencil point directed into the infundibulum and extending inferior to the chiasm (white arrowhead). In A, the sharp demarcation between the high signal intensity of the CSF of the suprasellar cistern and the low signal pulsation artifacts from the basilar artery also indicates the position of the membrane of Liliequist (see also Fig. 13-6B).
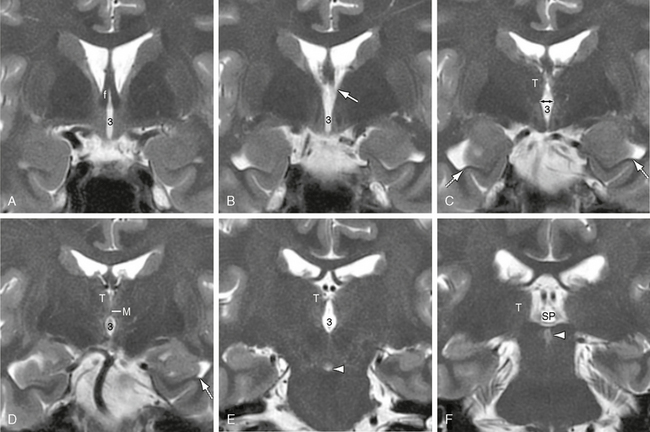
FIGURE 13-8 Third ventricle. A to F, Coronal T2W MR images. In order from anterior to posterior, coronal sections through the third ventricle (3) display the anterior columns of the fornices (f) that form the anterior walls of the foramina of Monro, the paired foramina themselves (white arrow in B), the diamond shape of the third ventricle where the hypothalamic sulci (double-headed black arrow in C) groove, the medial walls of the thalami (T), the constriction of the fluid space where the two thalami unite across the third ventricle at the massa intermedia (M), the re-expansion of the third ventricle behind the massa intermedia, and the posterior extension of the ventricle into the suprapineal recess (SP). The hypothalamic sulci mark the border between the thalami above and the hypothalami below. Also noted are the collateral eminences along the floors of the temporal horns (dotted white arrows in C and D), the interpeduncular fossa (arrowhead in E), and the aqueduct of Sylvius (arrowhead in F).
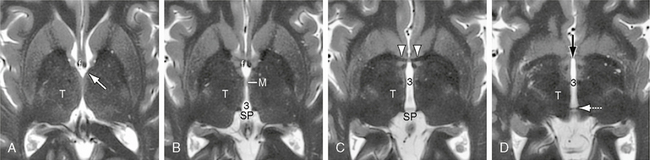
FIGURE 13-9 Third ventricle. A to D, Axial T2W MR images. In order from superior to inferior, axial images through the third ventricle (3) display the paired thalami (T), foramina of Monro (white arrow), massa intermedia (M), suprapineal recess (SP), the anterior commissure (white arrowheads) and very thin lamina terminalis (black arrow in D) that form the anterior wall of the third ventricle, and the habenular commissure (white dotted arrow) along the posterior wall of the ventricle. The lateral walls of the third ventricle are normally gently biconcave.
The third ventricle exhibits small extensions called recesses that project outward from the ventricle (see Figs. 13-2, 13-6, and 13-7). Anteroinferiorly, the supraoptic and infundibular recesses partially enclose the optic chiasm. The supraoptic recess lies between the lamina terminalis and the chiasm and points anteroinferiorly toward the cranial ends of the optic canals. The infundibular recess protrudes into the pituitary stalk (infundibulum) immediately posterior to the optic chiasm and points inferiorly to the pituitary gland. In coronal plane images, the supraoptic recess characteristically has the shape of an Erlenmeyer flask with its base resting on the chiasm. The infundibular recess characteristically has the shape of a long sharp pencil with its point directed inferiorly toward the pituitary gland.7,10 Behind the infundibular recess, a small midline hypothalamic recess extends inferiorly into the floor of the third ventricle.
Posterosuperiorly, the suprapineal recess passes posteriorly into the cistern of the vein of Galen above the pineal gland, and a small pineal recess passes posteriorly into the base of the pineal stalk (see Fig. 13-2). The suprapineal recess varies in length from 1 to 24 mm and in height from 2.5 to 9.0 mm.11 It is usually larger in young children and smaller in adults. In approximately 2% of patients, especially children, a large suprapineal recess is seen as a normal variation of no pathologic significance.9 In patients with severe hydrocephalus, the suprapineal recess may enlarge markedly and protrude downward through the incisura into the superior vermian cistern to compress the vermis or rupture into the subarachnoid space to create a free communication between the third ventricle and the quadrigeminal plate cistern (spontaneous ventriculocisternostomy).9
Cerebral Aqueduct
The aqueduct of Sylvius traverses the midbrain to interconnect the third ventricle with the fourth ventricle (see Figs. 13-2, 13-5H, and 13-10). In axial sections, the ventral wall of the aqueduct is pointed where it faces the tegmentum of the midbrain. The dorsal wall of the aqueduct is round where it faces the quadrigeminal plate. The aqueduct is considered in five segments. From anterior to posterior, these are designated the (1) adytum, (2) first constriction, (3) ampulla, (4) second constriction, and (5) egressus. The first and second constrictions correspond to impressions on the roof of the aqueduct by the overlying superior and inferior colliculi, respectively. The ampulla corresponds to the gap between those impressions at the level of the transverse intercollicular sulcus. The first constriction is normally the narrowest point along the aqueduct.12
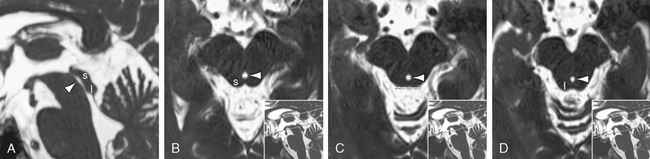
FIGURE 13-10 Cerebral aqueduct. T2W FIESTA MR images in the midsagittal plane (A) and an oblique coronal plane (B to D) perpendicular to the PC-O (posterior commissure-obex) line. In sagittal section, the ventral contour of the aqueduct (arrowheads) is smoothly concave along its length from superior to inferior. Anatomically, the dorsal contour shows narrowings at the level of the superior (s) and inferior (i) colliculi with a central expansion at the intercollicular level (dotted line). On imaging studies, the apparent caliber of the aqueduct is also affected by CSF pulsations. (See also Fig. 13-11E.)
Fourth Ventricle
The fourth ventricle lies in the middle of the posterior fossa (see Figs. 13-1 to 13-5 and 13-11). It connects rostrally with the third ventricle via the aqueduct of Sylvius. It connects caudally with the basal cisterns through three “exit foramina”: a midline foramen of Magendie that leads into the cisterna magna and paired lateral foramina of Luschka that lead into the lateral medullary cisterns (low cerebellopontine angle cisterns).13 The fourth ventricle is delimited anteriorly by the posterior surfaces of the pons and the upper medulla. Thus, the dorsal surface of the brain stem forms the floor of the fourth ventricle. This floor displays a median sulcus, paired paramedian sulci limitantes, transverse striae medullares, paired medial eminences and facial colliculi, and specific depressions (fovea) that mark out the positions of brain stem nuclei (see Chapter 15). The fourth ventricle is delimited anterosuperiorly by the superior vermis and superior (anterior) medullary velum. It is delimited posteroinferiorly by the inferior vermis and inferior (posterior) medullary velum. The fourth ventricle is delimited laterally by the superior and inferior cerebellar peduncles.
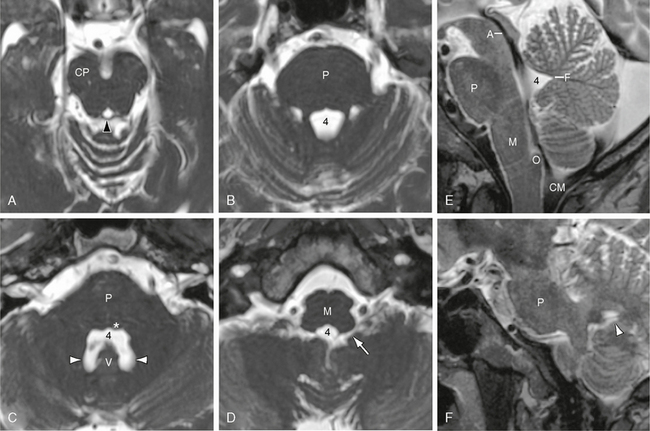
FIGURE 13-11 Fourth ventricle. T2W MR images in the axial (A to D) and sagittal (E and F) planes. A to D, Serial axial sections from superior to inferior show the fourth ventricle (4) expanding from its pointed “aqueductal” contour (black arrowhead in A) to the full “temple bell” configuration at its height (in C) (compare with Fig. 13-3C). E, Midsagittal images show the fastigial recess (F) most clearly. The paired posterior superior recesses (white arrowheads) are better appreciated in axial (C) and parasagittal (F) images. The lateral recesses (white arrow) pass anterolaterally from the fourth ventricle to the cerebellomedullary (lateral medullary) cisterns. In E, low signal flow voids depict the contour of the aqueduct (A) and the preferential flow of CSF from the aqueduct along the floor of the fourth ventricle to the midline foramen of Magendie and the cisterna magna (CM). The dorsal half of the fourth ventricle normally shows little flow effect. Also labeled are the cerebral peduncle (CP), medulla (M), obex (O), pons (P), vermis (V), and facial colliculus (asterisk in C) at the floor of the fourth ventricle.
Three recesses of the fourth ventricle project into the cerebellum posterosuperiorly (see Figs. 13-1, 13-3C, and 13-11). In the midline, the fastigial recess lies just above the nodulus at the junction of the superior and inferior medullary vela. To each side of midline, and slightly farther caudally, paired posterior superior recesses lie just above the superior poles of the tonsils. Laterally, paired lateral recesses lead from the floor of the fourth ventricle around the brain stem to the cerebellomedullary (lateral medullary) cisterns, where they end as the foramina of Luschka. At the caudal end of the fourth ventricle the paired inferior cerebellar peduncles converge to create an inferior point (the obex) that leads into the central canal of the spinal cord.
Cava Septi Pellucidi et Vergae
Cavum Septi Pellucidi
The septum pellucidum is a thin, vertically oriented structure situated in the midline between the two frontal horns (Figs. 13-12 to 13-14A to C). It is composed of two juxtaposed leaves of tissue. Each leaf is made of an ependymal-lined lateral surface, a thin layer of white matter, a thin layer of gray matter, and a medial (inner) pial layer.14 Developmentally, the two leaves arise separately and usually merge to form a 1- to 3-mm thick wall before 36 weeks of gestation.14 On each side, separately, each septal leaf stretches from the undersurface of the corpus callosum to the upper surface of the ipsilateral anterior column of the fornix. Persistent separation of the two septal leaves creates a midline CSF-filled space designated the cavum septi pellucidi. The cavum is delimited anteriorly and superiorly by the corpus callosum, laterally by each septal leaf, and inferiorly by the rostrum of the corpus callosum and the anterior columns of the fornices. It contains no choroid plexus.
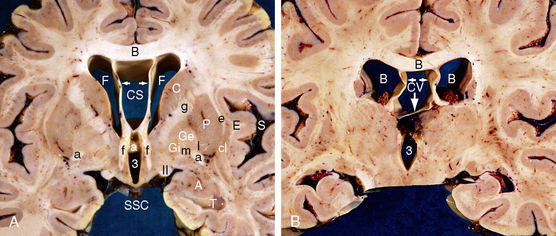
FIGURE 13-12 Large cavum septi pellucidi and cavum vergae. A, Cavum septi pellucidi (CS). Coronal anatomic section at the level of the anterior body of the corpus callosum (B), two frontal horns (F), cavum septi pellucidi (CS), anterior columns of the fornices (f), anterior commissure (white and black a) and anterior third ventricle (3). The paired leaves of the septum pellucidum (white arrows) extend vertically from the undersurface of the corpus callosum (black B) to insert onto the ipsilateral fornix (f). The cavum septi pellucidi lies between the corpus callosum above, the two septal leaves laterally, and the fornices inferiorly. The septal veins course within the septal leaves. The suprasellar cistern (SSC) lies inferiorly, the sylvian fissure (S) laterally and the temporal horn (T) and amygdala (A) inferiorly. Also labeled, from lateral to medial, are the extreme capsule (E), claustrum (cl), external capsule (e), putamen (P), lateral medullary lamina of the lenticular nucleus (l), external nucleus of the globus pallidus (Ge), medial medullary lamina (m) of the lenticular nucleus, internal nucleus of the globus pallidus (Gi), the optic tract (II), the caudate nucleus (C), and the genu (g) of the internal capsule. B, Cavum vergae (CV). Coronal anatomic section at the level of the posterior body of the corpus callosum (black B), bodies of the lateral ventricles (white Bs) and third ventricle (3). The cavum vergae (CV) lies inferior to the body of the corpus callosum, between the paired fornices (horizontal white arrows), and superior to the inferiorly displaced commissure of the fornices (vertical white arrow).
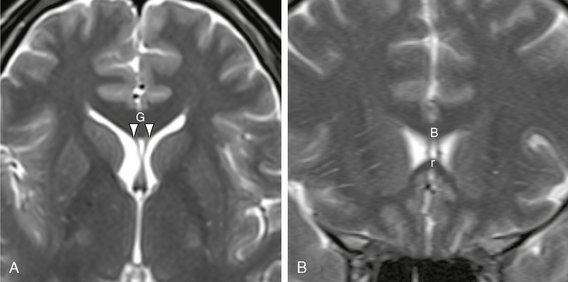
FIGURE 13-13 Small cavum septi pellucidi. Axial (A) and coronal (B) T2W MR images show a small triangular cavum septi pellucidi situated between the anterior aspects of the paired septal leaves (white arrowheads). The cavum septi pellucidi nestles within the curvature of the corpus callosum, delimited anteriorly by the genu (G), superiorly by the body (B), and inferiorly by the rostrum (r) of the corpus callosum.
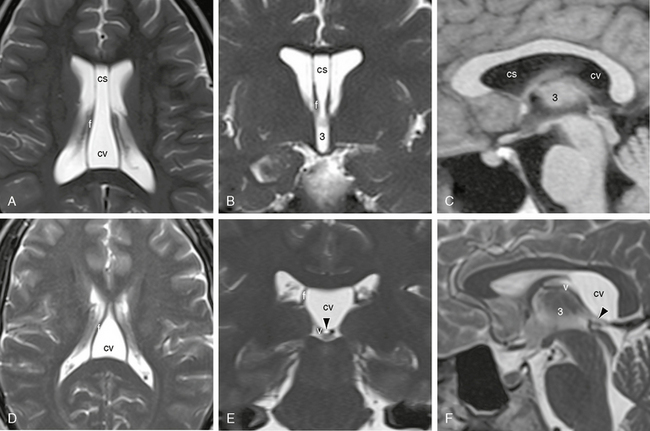
FIGURE 13-14 Cavum septi pellucidi (cs) and cavum vergae (cv). MR images. A to C, Combined cs and cv. Axial (A) and coronal (B) T2W MR images and sagittal T1W image (C) display the large midline cs between the paired septal leaves and direct extension of that cavity between the two fornices (f), indicating concurrent cv. The undersurface of the posterior body of the corpus callosum is slightly elevated and shows an expanded curvature. The roof of the third ventricle (3) is normal. D to F, Isolated cavum vergae (cv). Axial (D), coronal (E), and sagittal (F) T2W MR images show expansion of the CSF space between the two fornices (f), increased signal intensity within this space, and similar elevation and fullness of the undersurface of the corpus callosum. In E and F, a thin layer (arrowhead) separates the cv above from the compressed cistern of the velum interpositum below. The paired internal cerebral veins (v) are displaced inferiorly with the velum interpositum.
A cavum septi pellucidi is present in all premature infants/fetuses until 34 weeks’ gestation and shows decreasing incidence thereafter. In 108 cranial sonograms of normal newborns, a cavum septi pellucidi was present in 100% of neonates aged 24 to 34 weeks’ gestation, 69% at 36 weeks’ gestation, 54% at 38 weeks’ gestation, and 35% at 40 weeks’ gestation.15 The width, depth, and area of the cavum do not vary with gestational age.15 In adults, a cavum septi pellucidi is found in 2.0% to 2.5% of normal individuals. The incidence is reported to be higher in patients with mental retardation (15.3%), schizophrenia (23%), and trauma.16–19
Cavum Vergae
The paired fornices arch upward from each temporal lobe toward the midline, pass immediately anterior to the splenium and inferior to the body of the corpus callosum, and then arch downward behind the anterior commissure toward the mammillary bodies. Immediately below the posterior body of the corpus callosum, commissural fibers from each fornix cross to the other side, creating a thin flat sheet of commissural fibers designated the forniceal commissure (hippocampal commissure, psalterium). The upper surface of the forniceal commissure is closely applied to the undersurface of the corpus callosum but is not fused to it. Separation of the forniceal commissure from the undersurface of the corpus callosum creates a fluid space designated the cavum vergae (see Fig. 13-14). The cavum vergae is delimited superiorly by the undersurface of the corpus callosum, inferiorly by the upper surface of the forniceal commissure, and laterally by the paired alvei of the fornices. Expansion of the cavum displaces the hippocampal commissure inferiorly, thereby compressing the cistern of the velum interpositum and displacing the internal cerebral veins and posterior choroidal arteries inferiorly. The cavum vergae usually communicates anteriorly with a cavum septi pellucidi (see Fig. 13-14A to C) but may exist alone (see Fig. 13-14 D to F). It contains no choroid plexus.
Subarachnoid Cisterns
The subarachnoid space is the CSF-filled space situated between the outer arachnoid membrane (superficially) and the pia mater (deeply). The arteries, veins, and cranial nerves traverse these cisterns as they course between the skull and the brain.20
The subarachnoid fluid spaces form one confluent sea of fluid that extends over the complete surface of the brain (Figs. 13-15 to 13-17). For convenience, the spaces are classically divided into supratentorial (see Fig. 13-16) and infratentorial (see Fig. 13-17) compartments, with the understanding that these compartments interconnect with each other through the incisura. In both compartments, arachnoid septa and trabeculae partially subdivide the subarachnoid space into smaller, (in)constant compartments called cisterns.20 These cisterns may be single midline spaces or paired bilateral spaces. The arachnoid trabeculae that partition the cisterns have been given names to facilitate discussion. However, even the named supratentorial membranes are actually found at dissection in only 73% to 100% of hemispheres and are variably complete when found.20 For these reasons, the “boundaries” given for specific cisterns, and any indications that specific membranes “separate” one cistern from another, must be understood as general concepts, not as documentation of constant watertight barriers between isolated spaces.
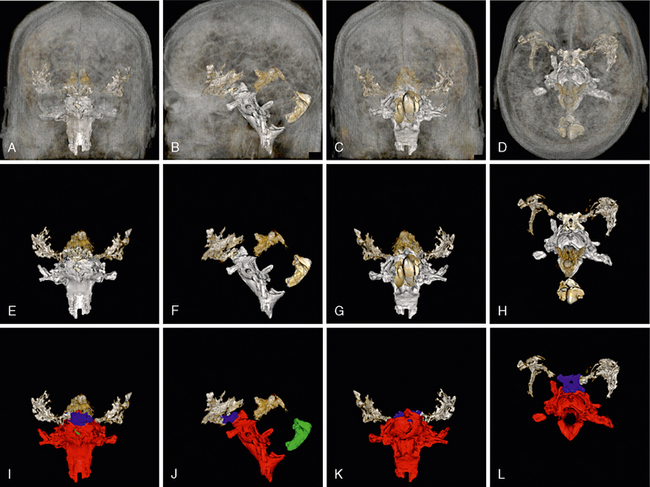
FIGURE 13-15 Overview of the subarachnoid cisterns. A to D, 3D reformatted MR images of the subarachnoid cisterns seen through a semitransparent scalp, skull, and brain. Anterior (A) lateral (B), posterior (C), and superior (D) views. E to H, Corresponding isolated subarachnoid cisterns oriented as A to D. I to L, Color coding of the cisterns displayed in A to H: silver, sylvian cisterns; blue, suprasellar cistern; red, the group of cisterns surrounding the brain stem; green
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


