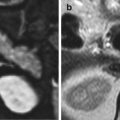
Fig. 1.1
(a) Anatomical coronal section of the brain showing the frontal lobes: peripheral cerebral cortex (arrow), white matter (star). (b) Axial section of the cervical spinal cord, coloured with the techniques for myelin. The grey matter (arrow) and the white matter (star) are highlighted
The CNS consists of two parts, anatomically connected through the foramen magnum, that are distinguished for their structures and positions: the brain, in the cranial cavity, and the spinal cord, in the spinal channel (see Chap. 2).
1.1.1 Meninges and Cephalorachidian Liquid
The CNS structures are enveloped in three layers of connectival membranes called meninges (the dura mater, the arachnoid mater, and the pia mater), located inside the osseous case (the skull and spinal canal of the spinal column; Fig. 1.2).
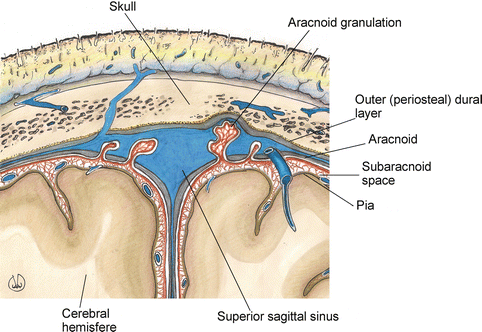
Fig. 1.2
Anatomy of the meningeal structures of the cerebral regions, coronal section
The dura mater is the most external part, it has a fibrous and resistant nature and perfectly adheres to the periosteum on the inner surface of the skull, from which it is separated by an anatomical space containing meningeal arteries and vessels (the epidural space). In the spine, the dura mater does not adhere to the spinal canal; rather, it is separated from it by the epidural space that contains adipose tissue and venous plexuses. The internal surface of the dura mater, in the endocranial part, is divided into three parts: the falx cerebri, which separates the two cerebral hemispheres; the tentorium cerebelli that exists between, and separates, the supratentorial region of the brain (telencephalon and diencephalon) from the subtentorial region (mesencephalon, pons, medulla oblongata, cerebellum), located in the posterior part; and the diaphragma sellae, a small stratum crossed by the hypophyseal peduncle.
The two layers of the dura mater contain vascular structures covered by endothelium tissue, named venous sinus, and they constitute the brain’s venous drainage system.
The arachnoid mater is the middle element of the meninges: it does not have vessels and it is similar to a spider web, containing the cephalorachidian liquid in the subarachnoid space (between the arachnoid and the pia mater). The arachnoid mater adheres to the dura mater’s internal surface, from which it is separated by the subdural cavity, which, in the skull area, is crossed by the “bridge” veins, while it is merely a virtual area in the spinal region. Since 10 years of age, the arachnoid mater is characterised by villi called arachnoid or Pacchioni’s granulations, located towards the venous sinus of the dura mater, that are involved in the reabsorption of the cephalorachidian liquid, helping the flow of the venous circulation (Fig. 1.2). The pia mater is the internal membrane, strictly adherent to the CNS surface; it contains a large amount of vessels continuing into the CNS. The pia mater of the brain, in correspondence with the brain’s ventricles, forms the tela chorioidea and the choroid plexus.
The dura mater and the spinal and brain’s arachnoid membrane follow one another in the foramen magnum region. The limit between the brain’s pia mater and the spinal pia mater coincides with the limit between the medulla oblongata and the spinal one. When exiting the brain cavities, at the level of the spinal canal, the three meninges envelop the brain and the spinal nerves, and continue to cover them.
The arachnoid mater and the pia mater are separated by the subarachnoid space in which the cephalorachidian liquid is contained; however, they are intensively connected to one another from the fibrous bridges of the subarachnoid space and therefore they are also considered to be a unique entity called leptomeninges. As the pia mater, contrary to the arachnoid mater, follows any prominence or depression of the neuraxis, the amplitude of the subarachnoid space is not regular, and presents wider regions at the brain level, called cisterns (base cisterns, chiasmatic cistern, Sylvian cistern, pericallosal cistern; Fig. 1.3).
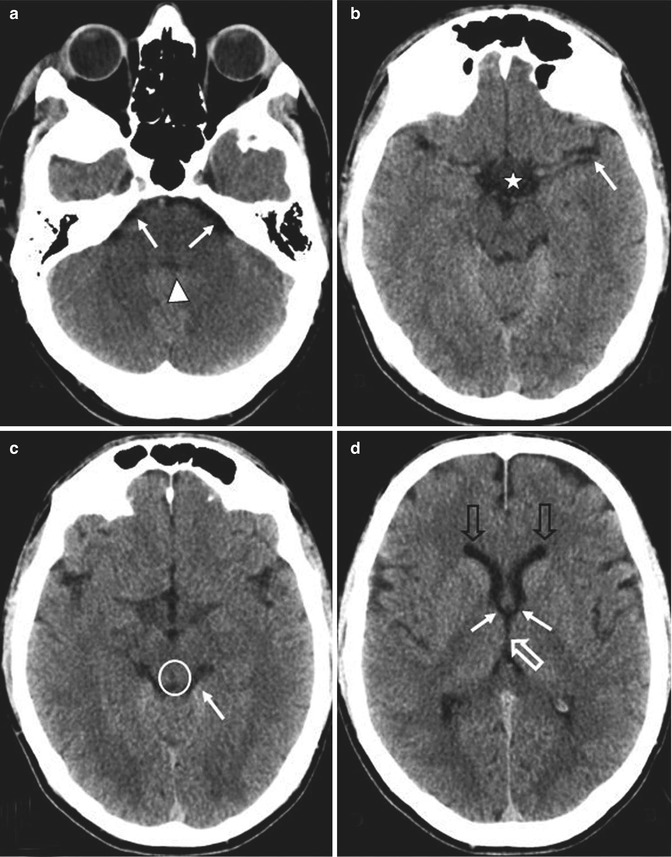
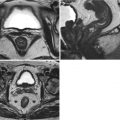
Fig. 1.3
Axial CT sections; ventricle and cistern fluid spaces. (a) Cerebellopontine angle cistern (arrows) and the fourth ventricle (arrowhead). (b) Sylvian fissure (arrow) and suprasellar cistern (star). (c) Quadrigeminal plate cistern (arrow) and aqueduct of Sylvius (in the circle). (d) Frontal horns of the lateral ventricles (empty black arrow), Monro foramina (arrows), third ventricle (empty white arrow)
The cephalorachidian liquid (whose function is to reduce the weight of the brain contained in it, to cushion it, and to enable the perfusion at a regular pressure) is produced at the level of the choroid plexuses (in the brain ventricles) and it is subject to its own dynamic movements, whose rhythm is provided by the cardiac rhythm. During the systole, the liquid from the lateral ventricles goes toward the third and fourth ventricles and, from there, into the subarachnoid spaces and the spinal canal. The opposite happens during diastole. Normally, the nervous system contains 150 ml of liquid, which is produced on a regular basis and needs to be regularly re-absorbed.
1.1.2 Brain
The brain, enveloped by the meninges, occupies the skull cavity. It is egg-shaped, with a sagittal major axis, weighing approximately 1,300 g in an adult person; the superior side is convex and the inferior side is flattened, the three posterior parts are rounded and they are called the frontal, temporal, and occipital lobes, the first are smaller, and located in the front part, the third one, at the back, is more voluminous (Fig. 1.4).
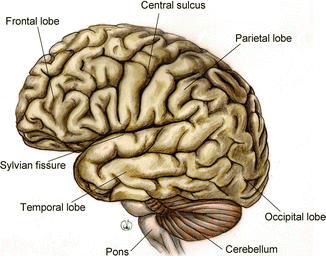
Fig. 1.4
Anatomy of the external cortical surface of the brain, sagittal section
From the macroscopic point of view, the brain is composed of an axial caudal part, the brainstem, and by the rostral super-axial centres that correspond to the voluminous masses of the telencephalon, above, and the cerebellum, behind.
The phylogenetical analysis shows that the brain is divided in three parts, derived from the primitive embryonic vesicle, the hindbrain, the midbrain, and the forebrain. The hindbrain is divided into the myelencephalon (medulla oblongata) and the metencephalon (pons or cerebellum); the midbrain remains as; the forebrain is divided into the diencephalon and telencephalon.
In the brain we find the ventricles, intercommunicating cavities where the cerebrospinal fluid flows (Fig. 1.5). The ventricular system, from the bottom to the top, consists of: the fourth ventricle, a cavity of the hindbrain – arriving from the central canal of the spinal cord, it is between the medulla oblongata and the pons – in the front, and the cerebellum, behind; the cerebral aqueduct of Sylvius, a thin cavity of the mesencephalon; the third ventricle, a cavity of the diencephalon; two lateral ventricles, cavities of the telencephalon, located in the relevant brain hemisphere and communicating with the third ventricle through the foramina of Monro.
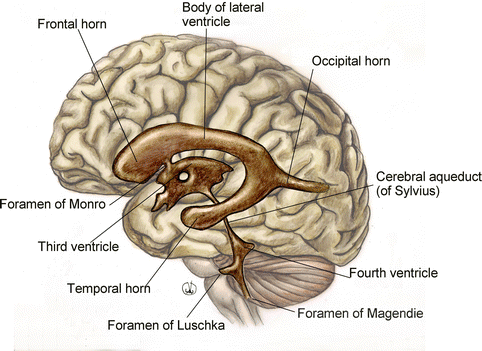
Fig. 1.5
Anatomy of the brain ventricular system
1.1.2.1 Brainstem
The brainstem is the axial part of the brain; it continues in the caudal area forming the spinal cord, at the level of the occipital foramen, and, upward, it continues into the diencephalon and the telencephalon. It is formed, from the bottom to the top, by the medulla oblongata, or bulb, the pons of Varolii, and the mesencephalon.
In the brainstem, we find the emergence of the cranial nerves, from the third to the twelfth, and the relevant nucleus, except for the spinal part of the spinal accessory nerve (ninth cranial nerve, CN9; Fig. 1.6).
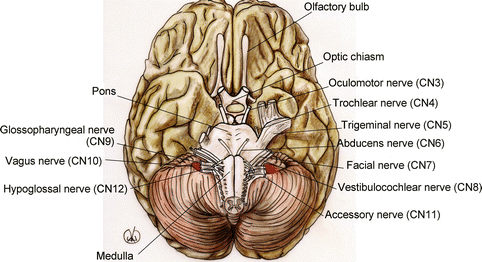
Fig. 1.6
Anatomy of the brainstem, with cranial nerves emergences (cn)
The medulla oblongata is located between the spinal cord, below, and the pons, above, and it is delimited by the bulbopontine sulcus. It is connected to the cerebellum through the inferior cerebellar peduncles. In the medulla oblongata, the motor nerves coming from the cerebral cortex, and going towards the spinal cord (corticospinal tract), form the pyramids that, in their inferior part, on the front side, cross each other and form the so-called pyramidal decussation.
The limit with the spinal cord is conventionally located in the space between where the last pair of cranial nerves emerge (twelfth, or hypoglossal nerve, CN12) and the first pair of cervical nerves.
The medulla oblongata also includes the nucleus responsible for the ocular reflexes of protection (eyelid reflex and tear production), of the superior respiratory reflexes (sneeze and cough), and feeding reflexes (suction and swallowing).
The pons is limited by the bulb, below, and the mesencephalon, above, that separate it, respectively, from the bulbopontine and the pontomesencephalic sulci. It looks like a ventral thickening in the brainstem and superficially it is streaked with transversal fibres, from one cerebral hemisphere to the other (the pontocerebellar fibres) that form on each side the middle cerebellar peduncles. On the front side of the pons, we find the basilar sulcus, which contains the basilar artery and the pontine cistern.
The mesencephalon is the portion of brainstem that connects the pons and the cerebellum to the diencephalon and the telencephalic hemispheres. It consists of the cerebral peduncles, in the front, and of the quadrigeminal plate, behind. The left and right cerebral peduncles are cylindrical trunks going outwards and upwards, delimiting the interpeduncular fossa, and, therefore, penetrating into the corresponding cerebral hemisphere. The quadrigeminal plate is a super axial centre that consists of four colliculi, two superior and two inferior, related to the visual and hearing system respectively, and connected to the lateral and the medial geniculate nucleus, which are diencephalic structures. The mesencephalon is also connected to the cerebellum through the superior cerebellar peduncles.
The internal organisation of the brainstem differs from that of the spinal cord with regard to the different location of the centres and of the pathways; the grey matter, in fact, is not gathered together, it is rather divided in smaller structures (cranial nerve nuclei and the typical nuclei of the brainstem) interposed with the ascending and descending ways: it is not therefore possible to clearly distinguish between white and grey regions, as it is in the spinal cord.
In the bulb, the pons, and the mesencephalon, we can distinguish between a ventral area, mainly occupied by white matter, and a dorsal area, where the grey matter is located (nuclei).
The brainstem regulates several essential functions; in particular, the reticular system (a group of neuronal populations that do not correspond to specific anatomical entities) plays an essential role, either directly or indirectly, in the control of the somatic motor functions, visceral motor functions or homeostasis (cardiac, vessels, respiratory, and visceral secretion functions), with the alternation of sleep–wakefulness, in the processes of addiction, adaptation to the environment, and transmission of general visceral afferents to the superior centres.
1.1.2.2 Cerebellum
The cerebellum is localised behind the medulla oblongata and the pons, and it is separated from them by the interposition of the fourth ventricle. It is situated in the posterior cranial fossa, contained in the cerebellar fossa of the squama occipitalis, and is separated by the tentorium cerebelli from the superior occipital lobes of the telencephalic hemisphere. The cerebellum has an elliptic shape with a large transverse axis, where there is a clear distinction between a central section – narrow and with a sagittal prolongation, which was called vermis by the first anatomists, because of the silkworm shape – and two lateral sections, most highly developed and corresponding to the cerebellar hemispheres.
On the bottom there is a depression, corresponding to the cerebellar hilus, where the superior, medial and inferior cerebellar peduncles join, and establish a connection with the mesencephalon, the pons and the medulla oblongata respectively. The cerebellum surface is transversely crossed by wide primary sulci, and we can distinguish between three lobes: anterior, posterior and flocculonodular. These lobes are divided into secondary sulci, consisting of thin circular convolutions called folia.
The cerebella tonsils are two lobules on the bottom side of the cerebellum, close to the bulb. They are on the lateral outline of the foramen magnum and may partly come out into the spinal canal. Their position is similar to that of the palatine tonsils, and this is the reason for their name.
A section of the brain shows a superficial mantle of grey matter, the cerebellar cortex, with a white matter underneath, named corpus medullare cerebelli, containing grey matter formations, corresponding to the intrinsic nuclei.
There are four cerebelli nuclei. On the medial axis, moving outwards: the fastigial, globose, emboliform and lateral or dentate. The first three nuclei are situated in the vermis, while the larger one, the dentate, is in the cerebellar hemisphere.
Systems of fibers reach the cerebellar cortex, bringing the stimuli coming from the peripheral system (from muscles, tendons, joints, sense organs). The cerebellar cortex is in fact a great integration centre for the stimuli that regulate and coordinate the muscular functions; the cerebellum, through the interposition of the ventral–lateral nucleus of the thalamus, sends impulses to the primary and secondary motor areas of the cerebral cortex to balance the voluntary movements. The cerebellum is also responsible for the body posture, ensuring the required tone. It also provides the required synergy and eumetria of the synergistic and antagonistic muscles, increasing the tone of the former, and decreasing the tone of the latter.
1.1.2.3 Diencephalon
The diencephalon is derived from the primitive vesicle of the forebrain, it is situated in the front end of the mesencephalon, and merges into it without interruption. It is laterally and dorsally surrounded by the cerebral hemispheres; the ventral section only is not coated by other nervous structures and, therefore, it can be seen from the outside. The median sagittal portion of the diencephalon consists of a thin fissure, the third ventricle, also called the diencephalic ventricle, which is characterised by a caudal narrowing that continues in the aqueduct of Sylvius. The existence, and the shape, of the third ventricle divide the diencephalon into two symmetrical parts, the right and the left. In turn, each of the said parts consists of a dorsal section, which is more substantial, and a ventral part, which is smaller.
The dorsal part of the diencephalon includes the dorsal thalamus (or thalamus proper) and a group of structures toward the dorsal–superior part of the same, forming the epithalamus.
The dorsal thalamus is the more substantial part of the diencephalon and the most important integration centre, intercalated by the sub-cortical structures and the cerebral cortex. All the different sensory functions, except for the olfactory one, convey into it before being projected to the cerebral cortex. The thalamus is therefore a sensitive station between the brainstem and cortex. The two thalami are connected through a bridge of grey matter called an intermediate or interthalamic commissure. The medial and lateral geniculate nuclei are linked to each thalamus. The lateral geniculate nucleus is larger and it is connected to the superior colliculus, while the medial geniculate nucleus is connected to the inferior colliculus. Retinal fibers convey into the lateral geniculate, from where the nerve fibers, projected into the visual cortex, originate; the medial geniculate is instead the commuting diencephalic station of the auditory pathway and its fibres establish a connection with the auditory cortex of the temporal lobe.
The epithalamus is a thin strip of tissue in the posterior–superior part of the dorsal diencephalon and it consists of the medullary stripe of the dorsal thalamus, the habenular trigone, the habenular commissure, the epiphysis (or pineal body) and the posterior commissure of the diencephalon. The ventral portion of the diencephalon is normally divided into the hypothalamus and the ventral thalamus (or subthalamus).
The hypothalamus is below the hypothalamic Monro sulcus and includes the plate of the third ventricle and the periventricular area (part of the base of the infundibulum, the tuber cinereum and mammillary bodies). The hypothalamus contains some of the most important integration centres of the autonomic nervous system and, therefore, takes part in the homeostatic regulation of some complex visceral functions (such as the body temperature), emotional expression, and the function of several endocrine glands. The hypothalamus, generally, has the important function of coordinating the endocrine glands, through a strong system of anatomical and functional connections with the hypophysis (hypothalamus–hypophyseal system). In the hypothalamus there are several groups of cells that constitute actual nuclei; among those, the paraventricular and supraoptic nuclei, which contain cells with secreting function (neurosecretion) and that produce oxytocin and vasopressin respectively, or antidiuretic hormone (ADH). These hormones are carried by the neurites, forming the supraoptic and the paraventricular nuclei, up to the neurohypophysis, where they are stored, to be released into the blood when the organism requires them. Through the hypophysis (or pituitary gland) the central nervous system in connected to the endocrine system.
The hypophysis consists of a glandular part, the adenohypophysis, embryologically derived from an invagination of the oral ectoderm (Rathke’s pouch), and a neural part derived from the diencephalon plate (neurohypophysis). Topographically, we can distinguish between a proximal tuberal part, smaller and located in the suprasellar area, and an intrasellar part, which is contained in a small bony cavity, the sella turcica, situated in the sphenoid bone, in the middle cranial fossa, at the base of the brain. The sella turcica is coated inside by the dura mater and it is covered by a dural fold, the diaphragma sellae, crossed by the pituitary stalk. The hypophysis has an ovoid shape, with a transverse diameter of approximately 1.5 cm, a sagittal diameter of 0.7–1.0 cm and a vertical diameter of 0.7–0.9 cm. In adult humans the hypophysis usually weighs less than 1 g, and it is normally bigger in men. The hypothalamus and hypophysis are closely connected through the hypothalamic–hypophyseal portal system, a set of vessels (Fig. 1.7). Inside such an anastomotic system, the hypothalamus, through the release of specific neurohormones (release factors or release-inhibiting factors), regulates the endocrine function of the hypophysis and of the organism.
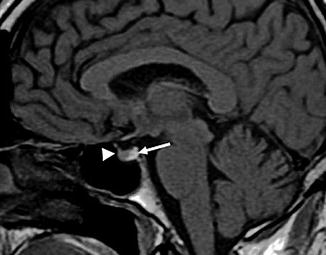
Fig. 1.7
MRI, sagittal T1-weighted image of the pituitary infundibulum. Adenohypophysis (arrowhead), isointense in comparison to the brain parenchyma, and neurohypophysis (arrow), hyperintense for the presence of vasopressin and oxytocin hormones, producing paramagnetic effects
The subthalamus (or ventral thalamus), that in caudal direction continues straight into the mesencephalic tegmentum, is on a side of the hypothalamus and below the grey matter of the dorsal thalamus. It forms an important subcortical station interposed with the pathways that regulate the activity of the voluntary muscles.
1.1.2.4 Telencephalon
The telencephalon originates from the telencephalic vesicle, which is the rostral part of the primitive proencephalic vesicle. It has an ovoid shape and occupies the largest part of the cranial cavity. The two hemispheres are separated by a deep fissure, the interhemispheric fissure, into which the falx cerebri penetrates; they are caudally separated from the brainstem by the transverse fissure.
Each hemisphere has three faces: the lateral, the medial, and the inferior, penetrating into each other corresponding to the margins, which may be more or less rounded. The lateral face is convex and connected to the internal surface of the cranial vault; the medial face is flat, vertically orientated, marking the hilus of the cerebral hemisphere, and therefore the area where the corpus callosum and the cerebral peduncle enter the cerebral hemisphere. The inferior face is connected to the cranial bones of the base, and, namely, from the front to the back, with the vaults of the orbits, the greater wings of the sphenoid bone, the sella turcica, the temporal bone pyramids, the occipital bone and the cerebellar tentorium.
Each hemisphere, from outside to inside, is composed of a thin layer of grey matter, the cerebral cortex or mantle; a central mass of white matter between the cortex and the basal nuclei; grey subcortical nuclei or basal nuclei (Fig. 1.8).
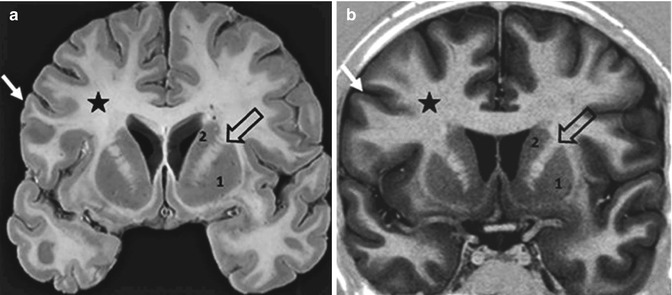
Fig. 1.8
Coronal anatomical section (a) and coronal MRI high resolution inversion recovery image (b). Cerebral cortex (arrow); white matter (star); basal nuclei: putamen (1) and caudate nucleus (2); internal capsule (empty arrow). The MRI images mostly correspond to the anatomical section
The external surface of the telencephalic hemispheres is characterised by several sulci and prominences that amplify their extension. They can be distinguished in primary fissures, dividing the hemispheres’ surface into lobes, and secondary fissures that cause in each lobe a series of circular convolutions or cerebral gyri.
Related posts:
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


