Fig. 2.1
Normal corpus callosum at 2 months (a), 1 year (b), and 14 years (c) on midsagittal T1-weighted images. White arrow anterior commissure, thick white arrows hippocampal commissure
The hippocampal commissure is part of the fornix, where about 20 % of the fibers cross the midline between the fornical crura, and form the transverse hippocampal commissure, a triangular transverse structure with an anterior vertex behind the septum pellucidum, a posterior base anterior to the forceps major, and a midline attachment to the undersurface of the callosal splenium [5]. The hippocampal commissure connects in humans the presubiculum and entorhinal and parahippocampal cortices, but not the hippocampus proper [4]. In an adult normal subject, its location can be identified on a sagittal MR section behind the callosal isthmus where the fornix joins the undersurface of the corpus callosum and in the concavity of the splenium [5] (Fig. 2.1).
The corpus callosum is the greatest forebrain commissure in placental mammals and specifically in humans. It has an interesting peculiar evolutionary history that parallels the striking increase of neopallium in the most developed mammals. It contains associative fibers arising from the vast majority of the cerebral hemispheres, and it is traditionally subdivided in five portions, the rostrum, the genu, the callosal body, the isthmus, and the splenium, that connect different cortical areas according to the following scheme [6]:
The rostrum is likely to connect the frontobasal cortex
The genu forms the anterior limit of the septum pellucidum. It connects the whole anterior frontal lobe, the prefrontal cortex, and the anterior cingulate area
The body connects the precentral cortex, the adjacent portion of the insula, and the overlying cingulate gyrus mostly.
The isthmus connects fibers of the motor and somatosensory strips and of the primary auditory area.
The splenium is the thickest part of the corpus callosum, and it connects the posterior parietal cortex, the medial occipital cortex, and the commissural fibers of the medial temporal cortex. As previously mentioned by its undersurface, the splenium is attached to the hippocampal commissure.
2.1 Commissure Development
The three commissures appear progressively from the 8 to the 13 GW.
2.1.1 Anterior Commissure
Between 7 and 8 GW, the anterosuperior wall of the future third ventricle is formed anteriorly by the lamina terminalis, in the region of the former closure of the cranial neuropore, and superiorly by the forming tela choroidea (Fig. 2.2a, b).
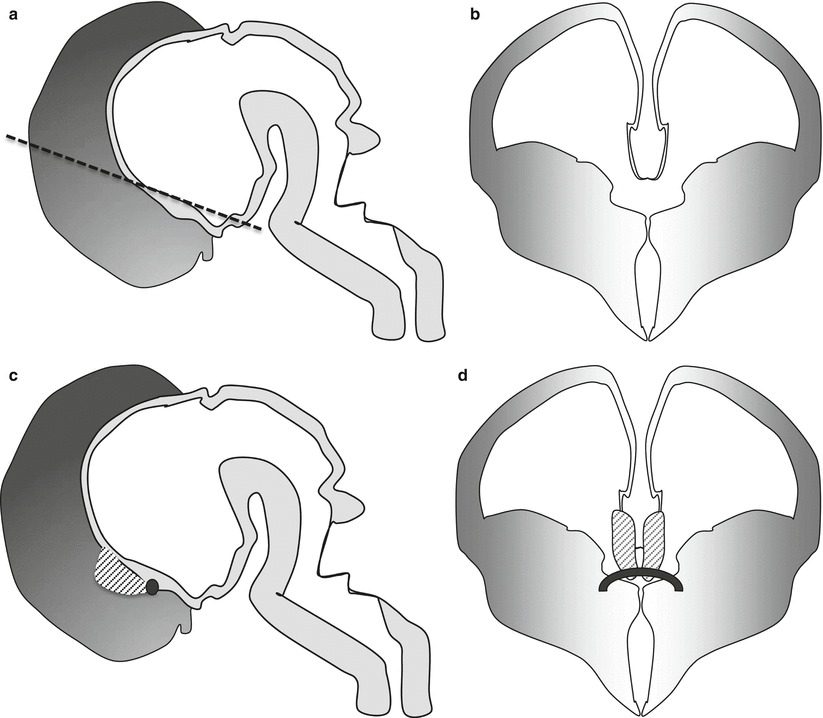
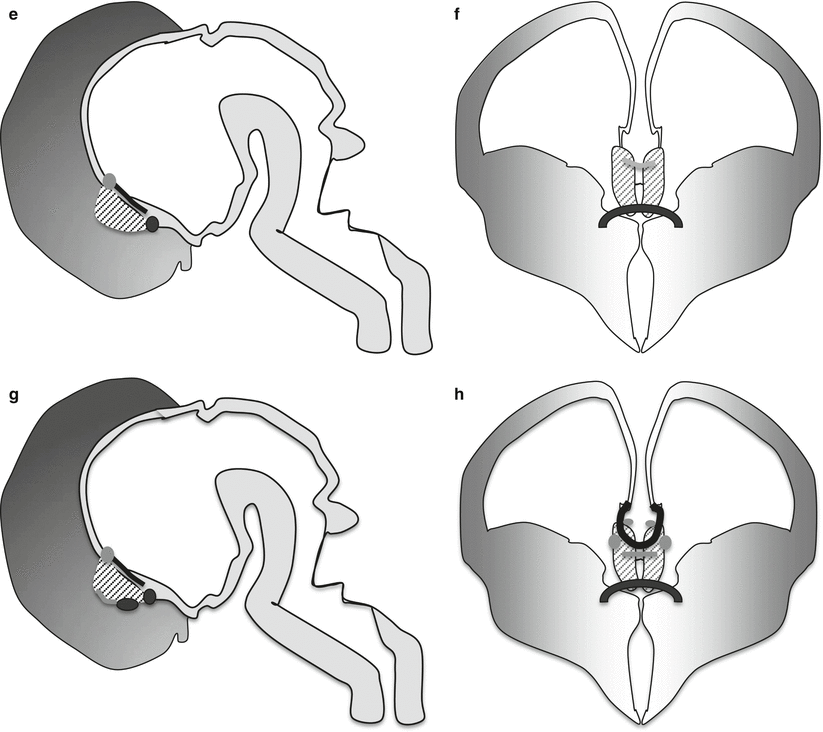
Fig. 2.2
Normal development of the three human commissures. Sagittal (a) and coronal (b) sections of the embryonic human brain at 7 GW, at this stage no commissures are still present. The anterior wall of the future third ventricle is formed by the lamina terminalis, whereas the superior wall is formed by the tela choroidea (the dotted line in sagittal section refers to the level of the coronal section and passes at the border of lamina terminalis an tela choroidea). Sagittal (c) and coronal (d) sections of the embryonic human brain at 8 GW. At this stage the lamina reuniens appears as a thickening of the upper and dorsal part of the lamina terminalis (dotted area). The first fibers of the anterior commissure appear at this stage in its inferior part (black circle c and black thick line d). Sagittal (e) and coronal (f) sections of the fetal human brain at 11 GW. At this stage the first crossing fiber of the hippocampal commissure appears at the dorsal part of the lamina reuniens (gray circle e, gray line f). The growing fornical fibers (black line in e) are in the inferior part of the lamina reuniens and define the inferior margin of future septum pellucidum. Sagittal (g) and coronal sections (h) of the fetal human brain at 13 GW. At this stage the pioneer axons of the forming corpus callosum (black oval g and curved black line h) cross the superior part of the SMTM guided by glial sling or midline zipper glia (small gray line h), glial wedge (gray ovals inferior and lateral to pioneer axons h), and indusium griseum glia (gray small circles superior and mesial to pioneer axons h) (drawings of this chapter by F. Triulzi)
During the 8 GW, the lamina reuniens becomes evident as a thickening of the portion of the lamina terminalis immediately adjacent to the tela choroidea [7], and it forms the shortest pathway between the hemispheres (Fig. 2.2c, d). The early fibers of the anterior commissure cross between the 9 and 10 GW in the lamina reuniens [7, 8].
2.1.2 Hippocampal Commissure
After the expansion of the fornical fibers during 9–11 GW, some of the fornical fibers start crossing the midline during 10–11 GW just in front of the attachment of the tela choroidea, where they form the early hippocampal commissure (Fig. 2.2 e, f). The crossing occurs either through the lamina reuniens itself [7] or at its surface, under the meningeal lining [8]. The segment of the fornix located between the septal nuclei and the hippocampal commissure represents the early septum pellucidum (Fig. 2.2 e, f).
2.1.3 Corpus Callosum
Around 11 GW a sulcus named sulcus medianus telencephali medii (SMTM) become evident in the region of the dorsal lamina reuniens [7]. The two lips of this sulcus come in close vicinity at the edge of the cortical plate [1], and glial cells and probably also neuronal cells [9] migrate medially from the subventricular zone toward the midline and invade the primitive meninx along the corticoseptal boundary to form a “glial sling” or “midline zipper glia” [5]; this process begins at about the 12 GW [7]. This interhemispheric glial bridge provides a support for the first pioneer axons to cross at about 12–13 GW [7], before disappearing shortly after [7]. The glial sling is independent anatomically and developmentally from both the anterior and the hippocampal commissures: it is located rostrally and dorsally to them [7].
Two other glial structures are needed to guide the pioneer callosal fibers toward the glial sling and the midline: the indusium griseum glia, located just above the corticoseptal boundary and the glial wedge located at the dorsomedial aspect of the lateral ventricle [10]. While the first axons begin to cross, the sling is still forming ventrally and dorsally.
At the end of the 13 GW, all the three commissures are eventually present, all in the close vicinity of the foramen of Monro: the anterior commissure within the ventral lamina reuniens, the hippocampal commissure within the dorsal lamina reuniens, and, rostral to the lamina reuniens, separated by the future septum pellucidum, the first corpus callosum pioneer axons from the cingulate areas (Fig. 2.2 g, h).
The corpus callosum (lamina rostralis, genu, and body fused with the splenium) is complete and easily recognized by the 14–15 GW [5, 11, 12], whereas the splenium becomes prominent by the 18–19 GW [11]. According to the more recent literature, the traditional theory that the corpus callosum would develop from the front to the back seems to be no more sustainable.
In humans due to the disproportionate expansion of the frontal lobes and consequently to the anterior accumulation of commissural fibers, the hippocampal commissure and its associated splenium are shifted posteriorly over the tela choroidea until the posterior third ventricle and the tectal plate.
This posterior translation of the splenium results from the anterior expansion of the frontal lobes and the corresponding accumulation of fibers in the anterior callosal segment and not from a front-to-back progression of the corpus callosum [5].
2.1.4 Corpus Callosum Development on Fetal MR and Fetal MR Autopsy
The shape of the corpus callosum is essentially similar to the final one by the 20 GW, and in a sagittal MR section, its anteroposterior extension is not far from what is usually seen in a newborn (Figs. 2.3 and 2.4); however, it is still extremely thin if it is compared with its fully developed stage [13]. Its final thickness is due to fiber addition during fetal life and for the greatest part by progression of myelination.
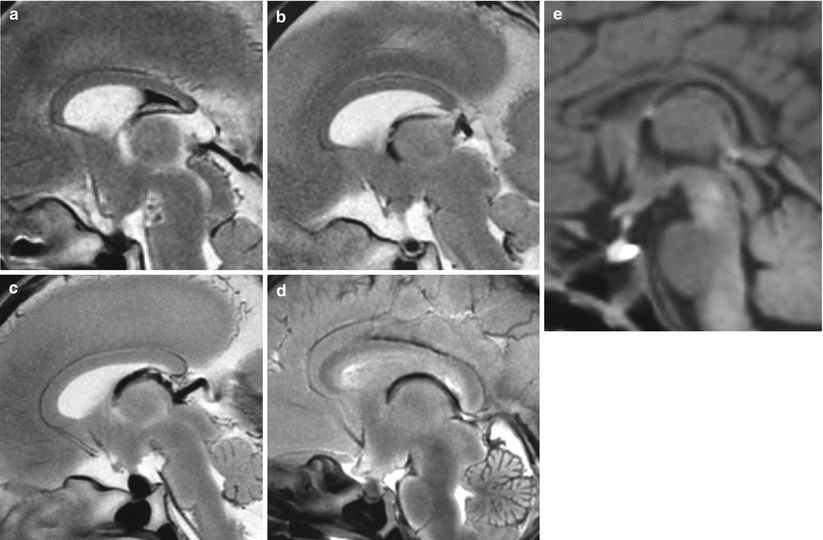
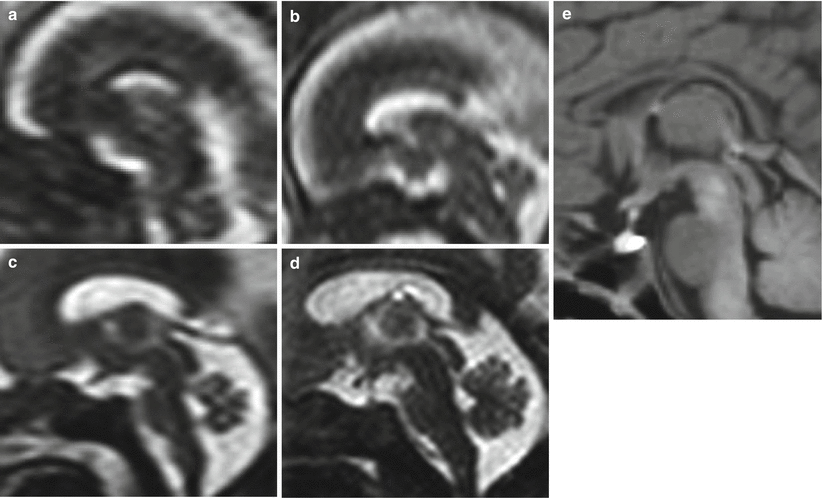
Fig. 2.3
Normal fetal development of corpus callosum on fetal MR autopsy midsagittal sections, (a) 19 GW, (b) 20 GW, (c) 22 GW, (d) 34 GW, (e) newborn at term. Even already visible in all its parts, the corpus callosum grows progressively in length from 19 GW to the term gestation. Comparing the 34 GW stage or the term newborn with the 19–20 GW stage, it is evident how the posterior part of the CC becomes progressively close to the pineal gland and to the quadrigeminal plate
Fig. 2.4
Normal fetal development of corpus callosum on fetal MR, midsagittal sections, (a) 19 GW, (b) 20 GW, (c) 22 GW, (d) 34 GW, (e) newborn at term. A particular care should be taken in the evaluation of CC morphology at 18–19 GW due to its reduced length in comparison with older fetuses
2.2 Commissure Malformations
2.2.1 Complete Commissural Agenesis
In the classic complete commissural agenesis, both the fibers of the corpus callosum and of the hippocampal commissure are actually present but do not cross the midline rather they form the parasagittal bundles of Probst for the corpus callosum and the lower edge of the medial telencephalic medullary velum together with the ipsilateral fibers of the fornix for the hippocampal commissural fibers; the accumulation of the callosal and fornical fibers explains the thick appearance of the velum.
In about 50 % of the cases, the anterior commissure is either absent or too thin to be recognized [5]; this could be due to the absence of its neocortical component (Figs. 2.5 and 2.6). The interhemispheric fissure is wide being the tela choroidea of the third ventricle the only structure that bridges the hemispheres. The lateral ventricles are away from the midline and parallel to each other, with thigh frontal horns medially stretched by the bundles of Probst and large occipital horns. The enlargement of the occipital horns (Figs. 2.5 c, g and 2.6 d, f, i) could be probably due to the absence/reduction of the forceps major [14] even if this hypothesis has not been definitely confirmed. In a recent review of 67 cases of complete CC agenesis studied by fetal MR the complete agenesis of all the three commissures resulted to be a very rare condition. In particular the anterior commisure could be detected in more than 90 % of cases and the hippocampal commissure in more than 50% of cases (Fig. 2.5, 2.6, 2.8, 2.9) [15].
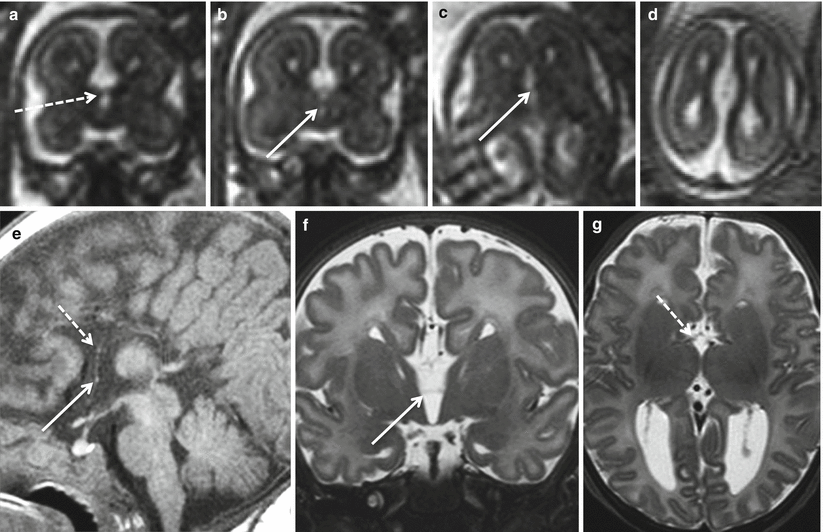

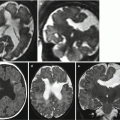
Fig. 2.5
Complete agenesis of the corpus callosum. Fetal MR study at 22 GW (a–d) and MR study in newborn at term (e–g). Only a thin anterior commissure (white arrows) is visible on both fetal (b, c) and neonatal studies (e, f). A doubtful minimal thickness of the region between lamina terminalis and tela choroidea is visible in both fetus (dotted arrow a) and in newborn (dotted arrow e, g)
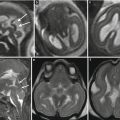
Fig. 2.6
Complete agenesis of the corpus callosum. Fetal MR study at 21.6 GW (a–c), 33 GW (d–e) and MR study in newborn at term (g–i). In this case the anterior commissure (white arrow) is well evident in the 33 GW MR fetal study (d) and in the post natal MR study (g, h). A minimal thickness of the region between lamina terminalis and tela choroidea (dotted arrow) is well visible in both fetal MR studies (b, e), whereas it appears more difficult to demonstrate in the post natal MR study (g, i)
2.2.2 Partial Commissural Agenesis
According to Raybaud [5], from an analysis of the commissural malformations based on the modern embryological understanding, partial agenesis is not different degrees of a continuum but rather different malformations, each with a specific developmental disorder and, potentially, a specific genetic defect.
Figure 2.7 summarized four different potential conditions of commissural abnormalities from a complete absence of the CC to partial hypogenesis. In A only the anterior commissure is evident, and in B a remnant of the hippocampal commissure is present in its original starting position, whereas in C an extremely small hypogenetic CC is present just anteriorly and superiorly with respect to the hippocampal commissure. Finally in D the CC is clearly present as well as the septum pellucidum comprises between its inferior aspect and the fornices.
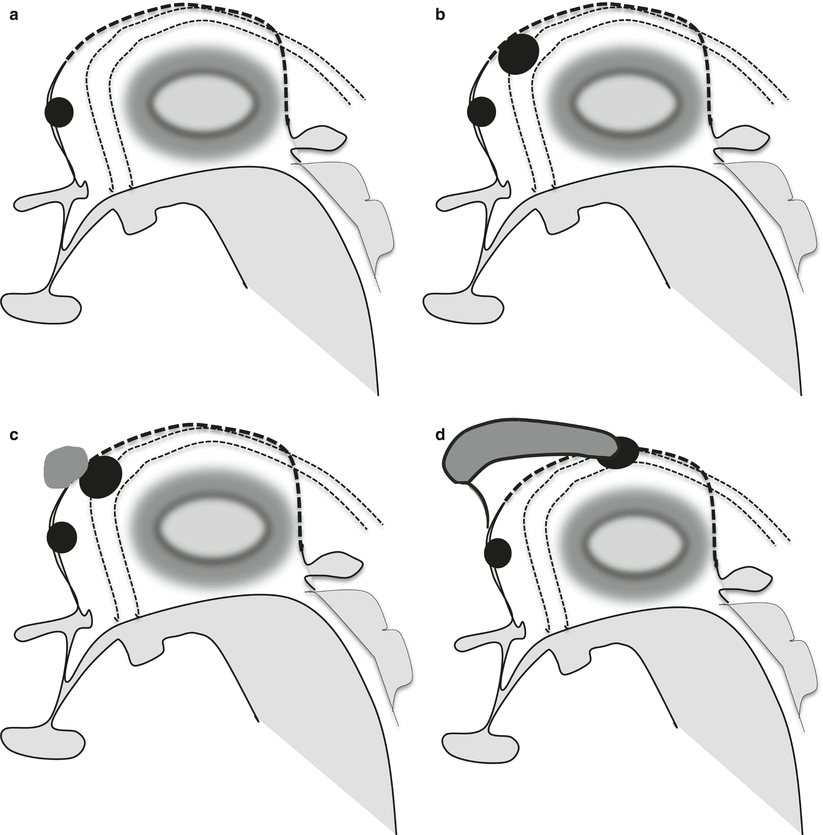
Fig. 2.7
Partial commissural agenesis. A complete commissural agenesis is extremely rare, in the real complete agenesis of the corpus callosum the anterior commissure (black oval in the superior part of the lamina terminalis) is usually present (a). Sometimes not only the anterior commissure is present but also a small hippocampal commissure is visible on midsagittal section (b) (large black oval between the lamina terminalis and the tela choroidea, dotted line, centered between the forniceal columns, double dotted lines). In agenesis of corpus callosum an extremely small remnant of CC can be detected just anterior-superiorly to the hippocampal commissure (c) (gray oval anterior to the hippocampal commissure). In the more typical CC partial agenesis the CC is partly recognized and the hippocampal commissure is pushed posteriorly (d), the septum pellucidum becomes visible between the fornices and the inferior part of the CC
Such different malformations can be detected also in fetal and neonatal MR studies. In Fig. 2.5, 2.6 and 2.8 a residual hippocampal commissure seems to be detectable in both fetal and neonatal studies. In Fig. 2.9 the hippocampal commissure seems to be larger and well evident. In Figs. 2.10 and 2.11 a small CC can be detected, whereas in Fig. 2.12 the progressive development of an hypogenetic CC can be followed from the 22 GW up to 3 years of age.
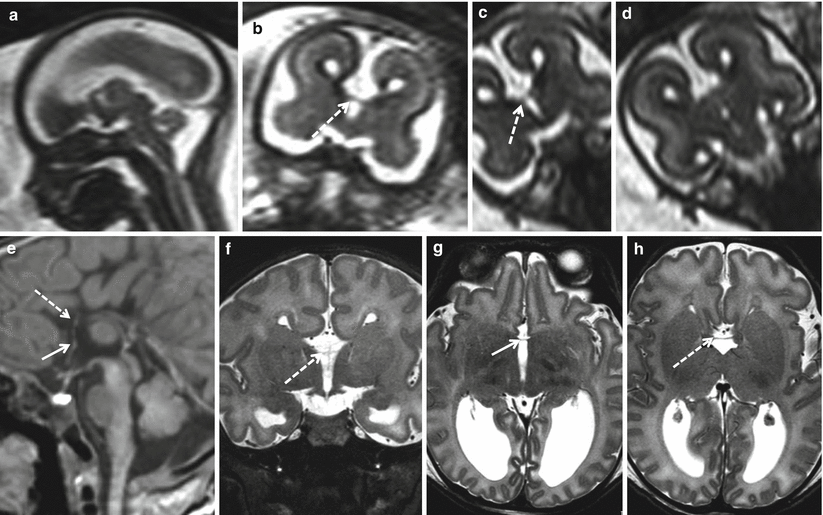
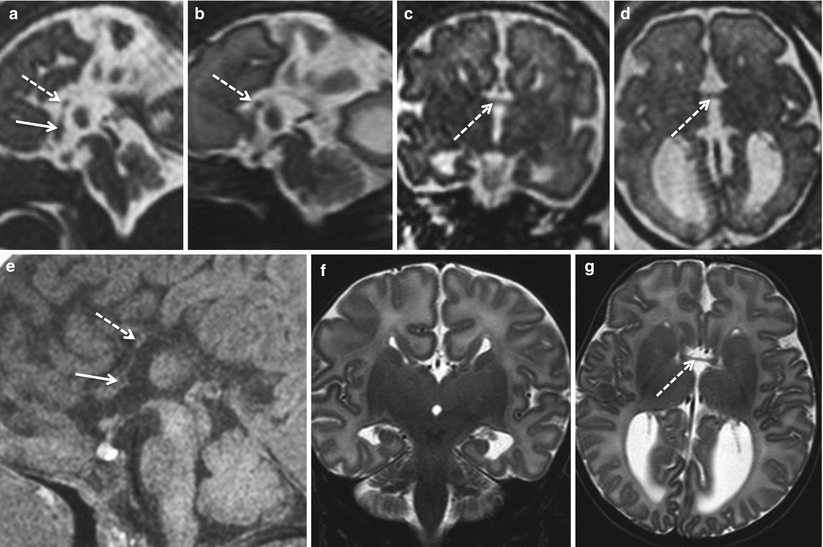
Fig. 2.8
Partial commissural agenesis. Fetal MR study at 20.6 GW (a–d) and MR study in newborn at term (e–h). A relatively thin hippocampal commissure is visible in both fetal and neonatal MR studies (dotted arrows). An extremely thin anterior commissure seems visible only in post natal MR (white arrows)
Fig. 2.9




Partial commissural agenesis. Fetal MR study at 31 GW (a–d) and MR study in newborn at term (e–g). The anterior commissure is barely visible (white arrows), whereas the hippocampal commissure is well evident even on the fetal MR study (white dotted arrows). Hippocampal commissure can be differentiated from a marked hypogenetic CC by its position that should be more inferior and posterior, bridging the fornices at the passage between lamina terminalis and tela choroidea. In this case however the presence of an extremely hypogenetic CC close to hippocampal commissure cannot be totally excluded
Related posts:
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
