FIG. 19.1 Ventral view of the cranial nerves at the base of the brain. The 12 paired cranial nerves are numbered from cranial to caudal in order of their attachment to the forebrain (CN I and II), midbrain (CN III and IV), pons (CN V to VIII), medulla (CN IX, X, and XII), or upper cervical spinal cord (CN XI).
(From Brain Anatomical Chart. Baltimore: Anatomical Chart Company, Wolters Kluwer Health and Pharma, 2000.)
The cranial nerves can also be organized by function. The olfactory (I), optic (II), and vestibulocochlear (VIII) nerves are sensory nerves, while the oculomotor (III), trochlear (IV), abducens (VI), spinal accessory (XI), and hypoglossal (XII) nerves are motor nerves. The trigeminal (V), facial, glossopharyngeal, and vagus (X) have both motor and sensory components. The functions of the cranial nerves are commonly annotated with letter symbols to indicate whether they innervate voluntary or involuntary muscles and/or receive general or special sensory information (Table 19.1). In the brainstem, the cranial nerve nuclei are organized from cranial to caudal as would be expected based on their attachment sites to the surface of the brain. However, nuclei of common functional type are also organized from medial to lateral, with motor “columns” lying medially and sensory “columns” lying more laterally (Fig. 19.2).
Table 19.1 Functional Components of the Cranial Nerves
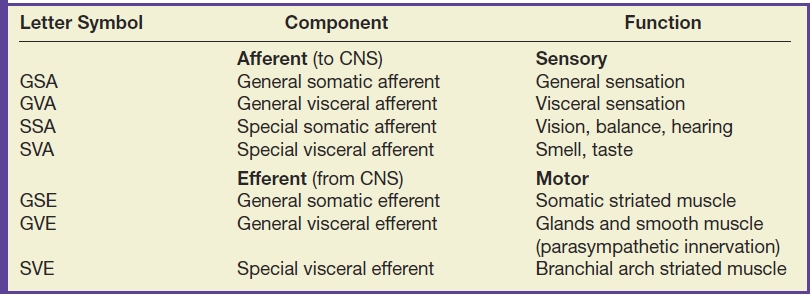
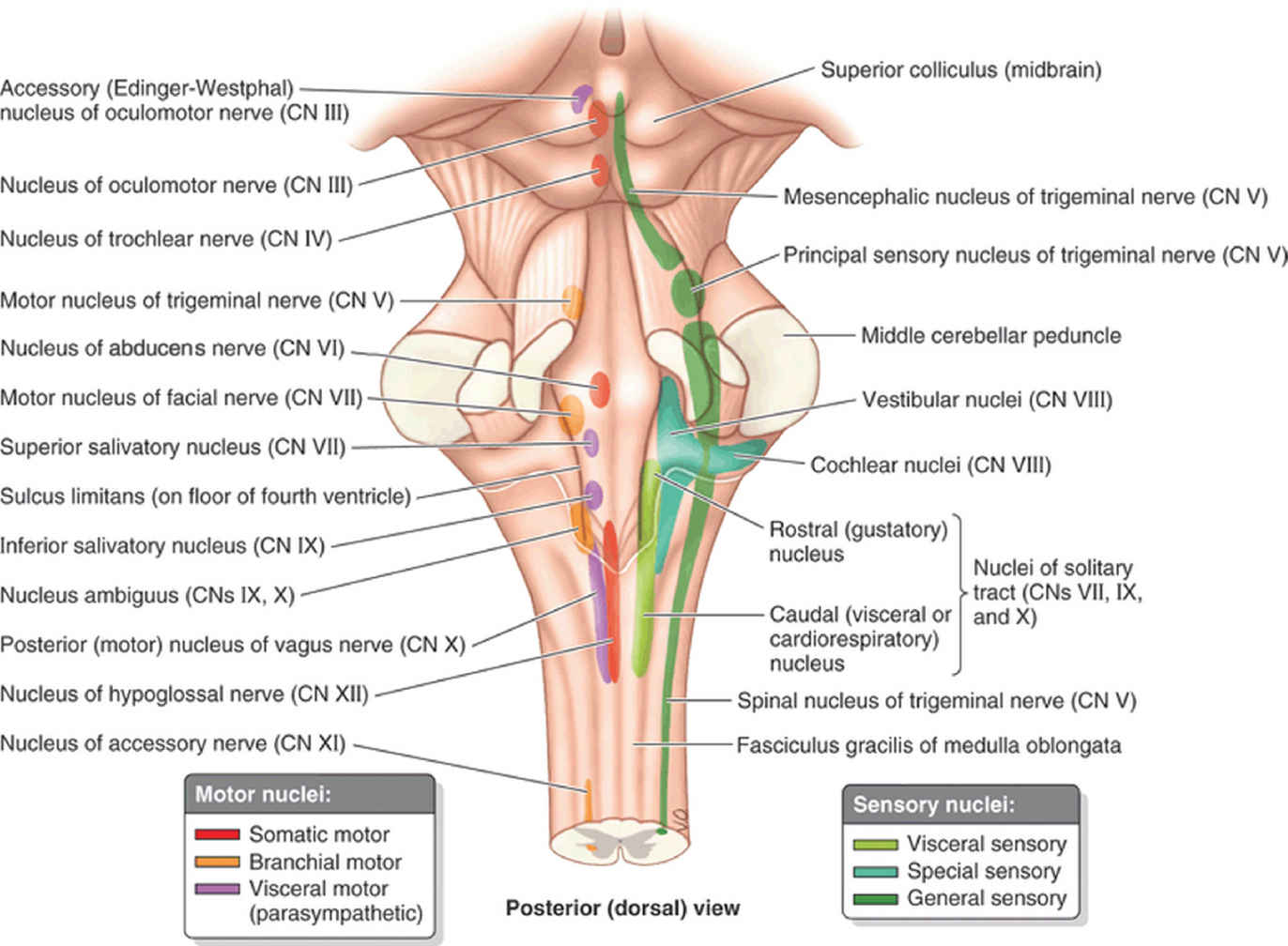
FIG. 19.2 Cranial nerve brainstem nuclei. The brainstem nuclei of the cranial nerves are organized in columns, with the motor nuclei lying closer to the midline and sensory nuclei located more laterally.
(From Moore KL, Agur AM, Dalley AF. Clinically Oriented Anatomy. 7th ed. Philadelphia, PA: Lippincott Williams & Wilkins; 2013.)
The cranial nerves, their motor and sensory components, functions, attachment sites to the brain, and associated skull base foramina and fissures are summarized in Table 19.2. The individual cranial nerves and their normal appearance at imaging are discussed below.
Table 19.2 Summary of the Cranial Nerves
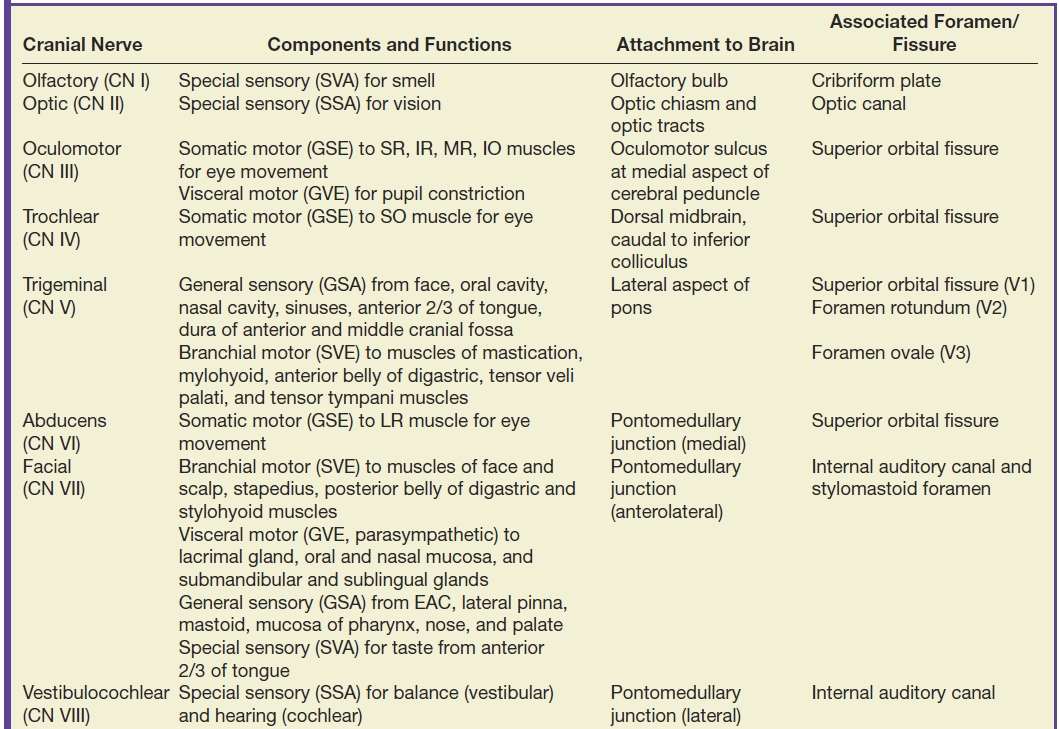
EAC, external auditory canal; GSA, general somatic afferent; GSE, general somatic efferent; GVA, general visceral afferent; GVE, general visceral efferent; IO, inferior oblique; IR, inferior rectus; LR, lateral rectus; MR, medial rectus; SO, superior oblique; SR, superior rectus; SSA, special somatic afferent; SVA, special visceral afferent; SVE, special visceral efferent; TM, tympanic membrane.
Olfactory nerve (CN I)
The intracranial olfactory bulbs and tracts of CN I are in actuality tracts of the central nervous system (CNS) rather than true peripheral cranial nerves. The olfactory bulbs and tracts lie along the cribriform plate, just inferior to the olfactory sulcus, which is located between the gyrus rectus and medial orbital gyrus. The primary sensory neurons of olfaction are actually located in the olfactory epithelium in the upper nasal cavity, which send their axons through the foramina of the cribriform plate to synapse on the olfactory bulbs. The olfactory nerves are best evaluated with coronal high-resolution MR imaging while the integrity of the cribriform plate is best assessed with coronal reformatted CT images of the anterior skull base (Fig. 19.3).
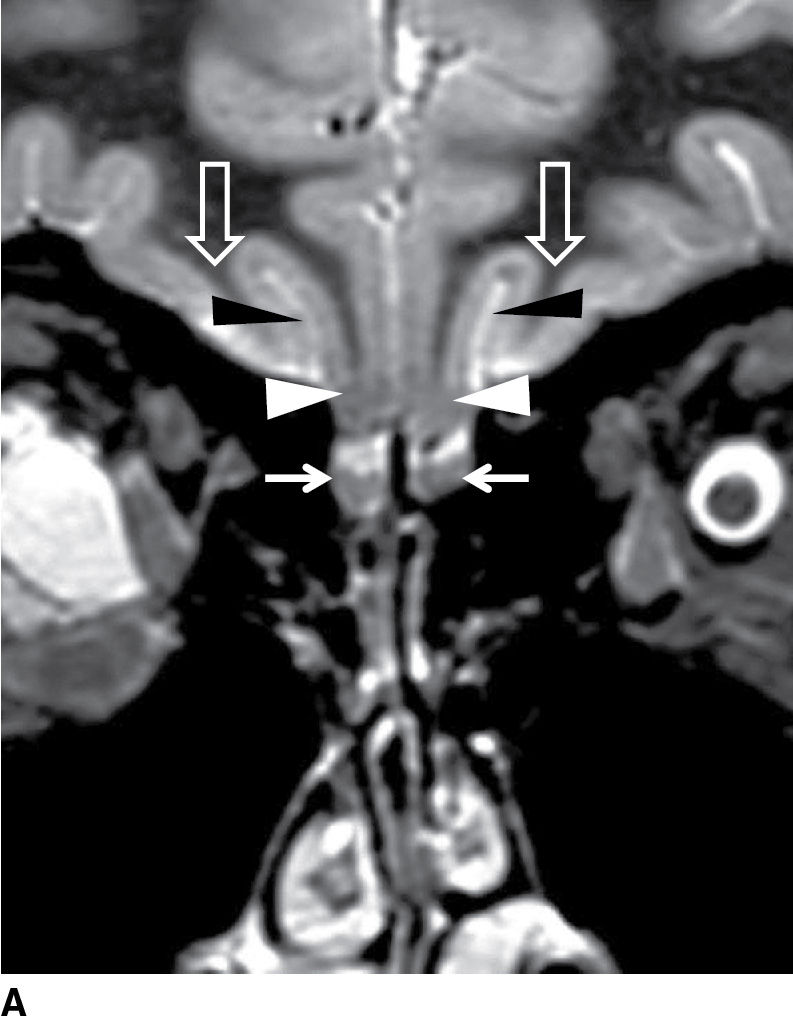
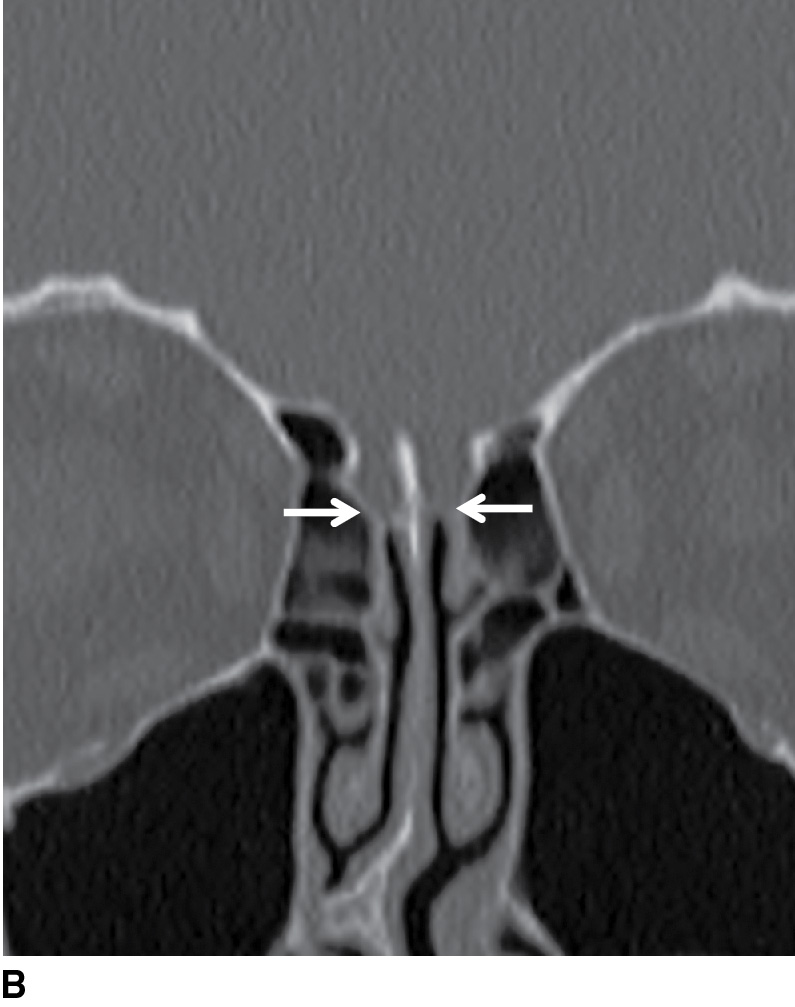
FIG. 19.3 Normal imaging appearance of the olfactory nerves. A: Coronal high-resolution T2-weighted image demonstrates the normal location and appearance of the olfactory tracts (thin white arrows) at the base of the anterior cranial fossa. The olfactory sulci (black arrowheads) are located just superior to the olfactory tracts, between the gyri recti (white arrowheads) and the medial orbital gyri (open arrows). B: The thin cribriform plate (arrows) can be used as a marker for the location of the olfactory tracts on coronal reformatted CT imaging.
Optic nerve (CN II)
The optic nerves, like the olfactory nerves, are not true peripheral cranial nerves but are instead extensions of the CNS. The primary sensory cells for vision are located within the retina, with the axons of the retinal ganglion cells forming the intraocular optic nerves (also called the optic nerve heads), which are normally not well seen at imaging. The radiologist can evaluate the remaining segments of the optic nerves, namely, the intraorbital (between the globe and the orbital apex), intracanalicular (within the optic canals), and intracranial (traversing the CSF space between the optic canals and the optic chiasm) segments (Fig. 19.4A–C). At the optic chiasm, fibers from the temporal aspects of the globes pass to the optic tract without crossing the midline while the nasal retinal fibers decussate and continue in the contralateral optic tracts. This explains the “x-shaped” configuration of the optic chiasm as seen on axial images (Fig. 19.4D). A detailed discussion of the optic pathway is beyond the scope of this chapter; however, in patients with visual field deficits, knowledge of the organization of these fibers helps predict the site of pathology. The structures of the optic pathway and their corresponding visual field defects are summarized in Figure. 19.5.
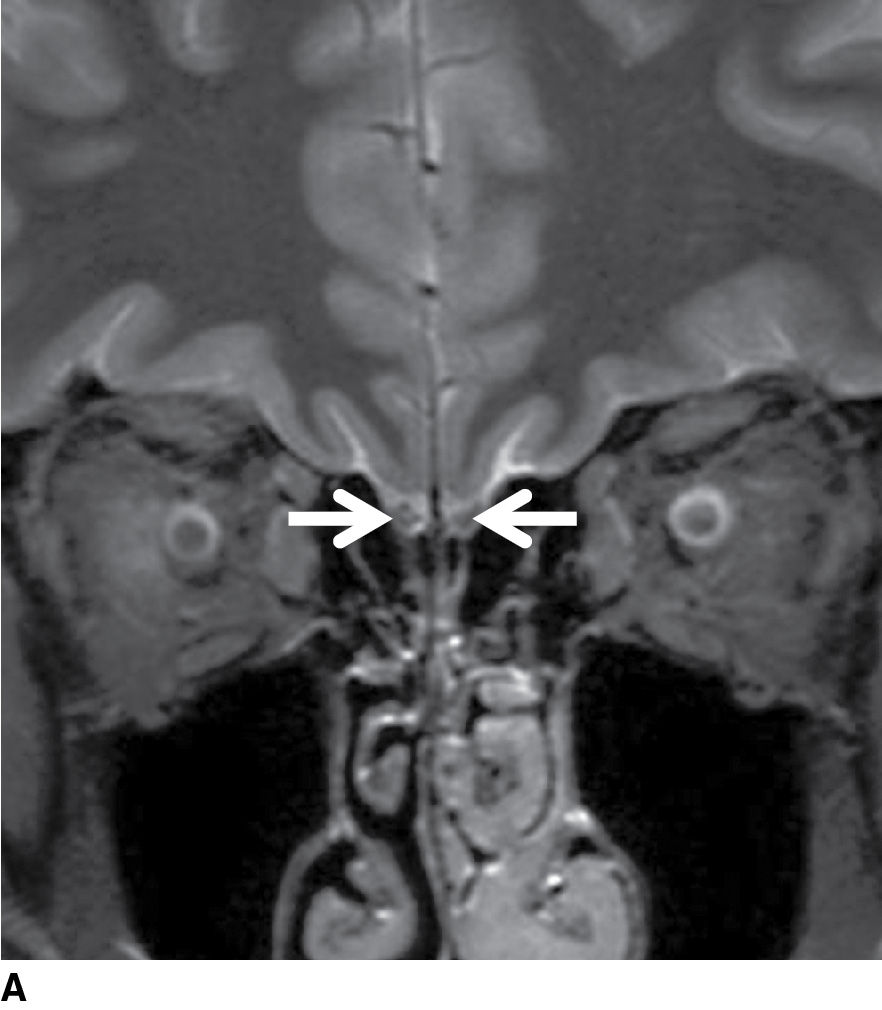
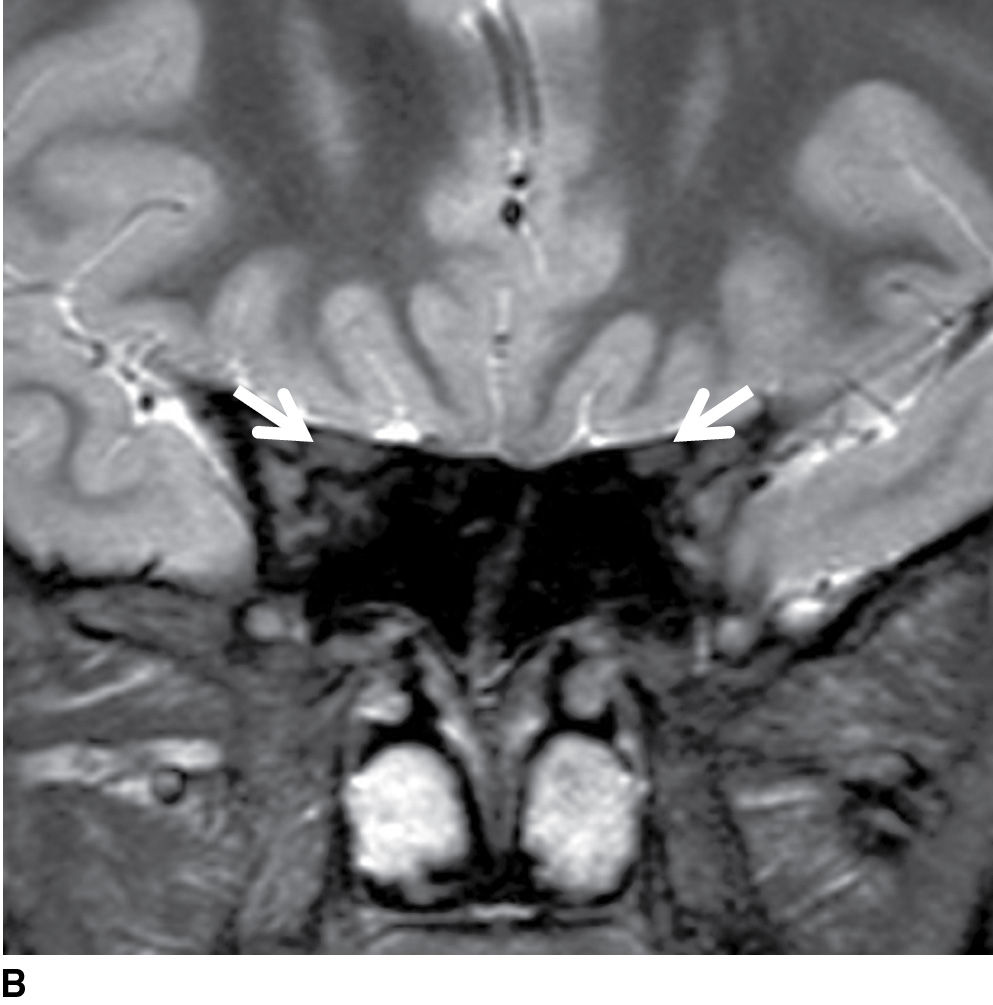
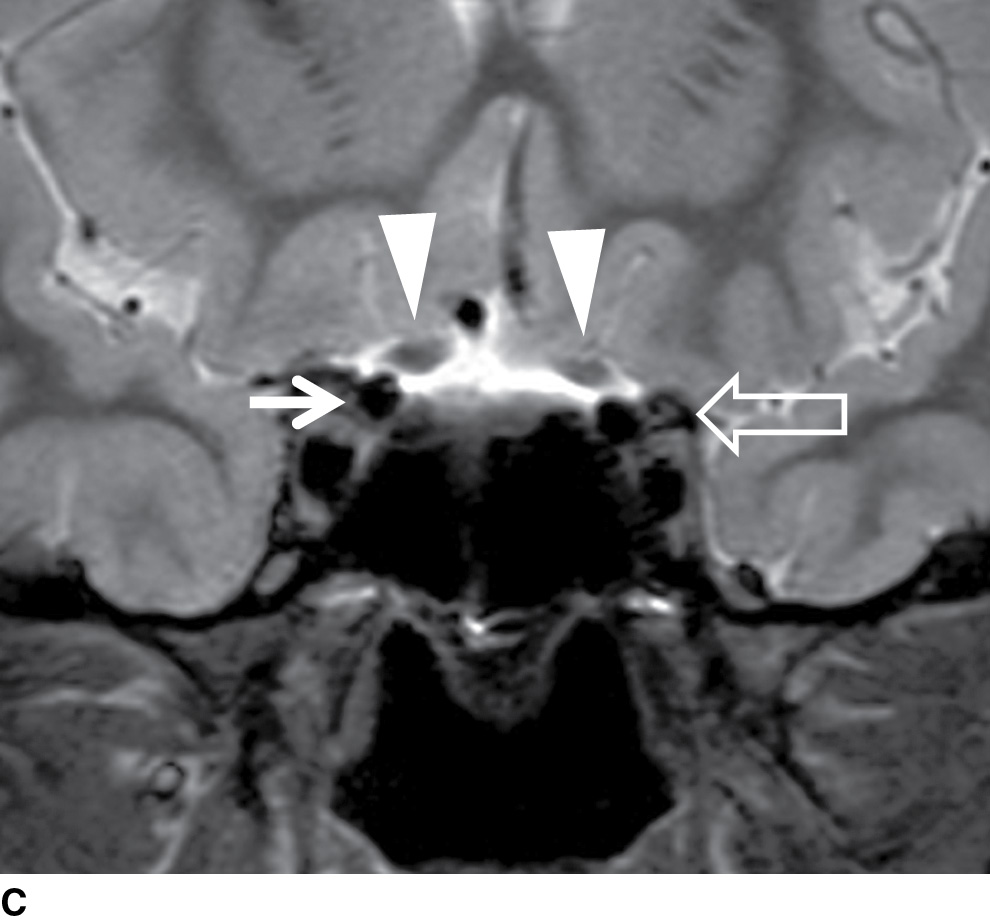
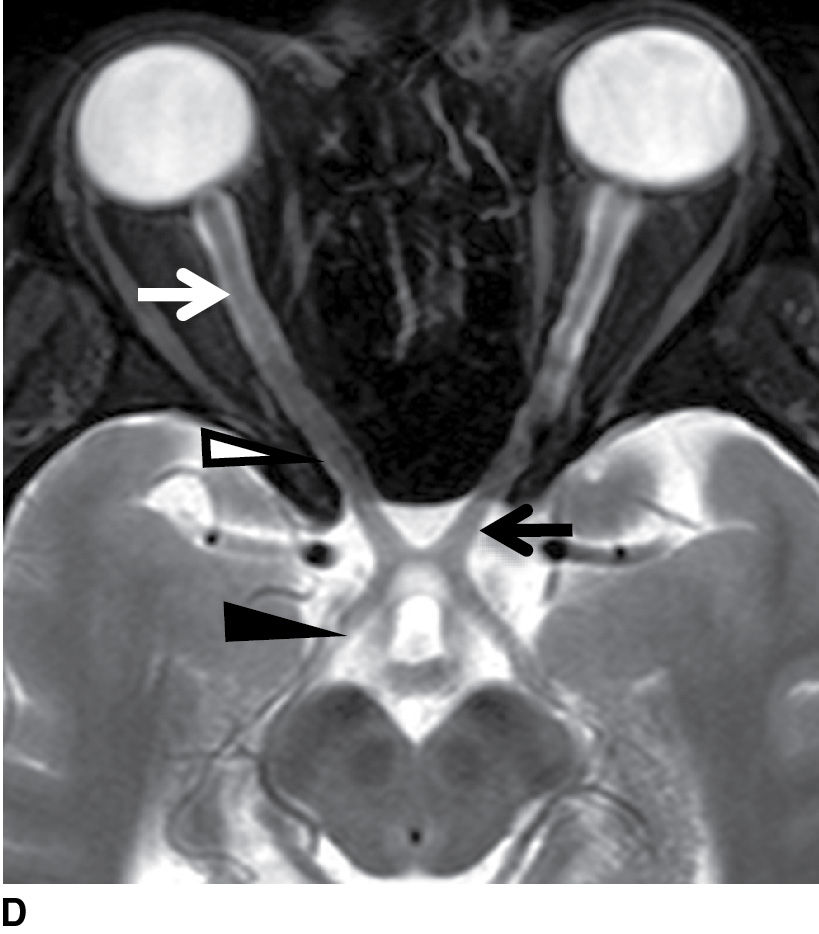
FIG. 19.4 Normal imaging appearance of the optic nerves and chiasm. Coronal STIR images demonstrate the normal appearance of the intraorbital (white arrows in A), intracanalicular (white arrows in B), and intracranial segments of the optic nerves (white arrowheads in C). The signal in the optic nerves matches that of the white matter in the brain. C: The intracranial optic nerves (arrowheads) lie superior to the internal carotid arteries (thin white arrow) and medial to the anterior clinoid processes (open arrow). D: The optic nerves in their entirety are seen on the axial T2-weighted image. White arrow, intraorbital optic nerve; white arrowhead, intracanalicular optic nerve; black arrow, intracranial optic nerve; black arrowhead, optic tract.
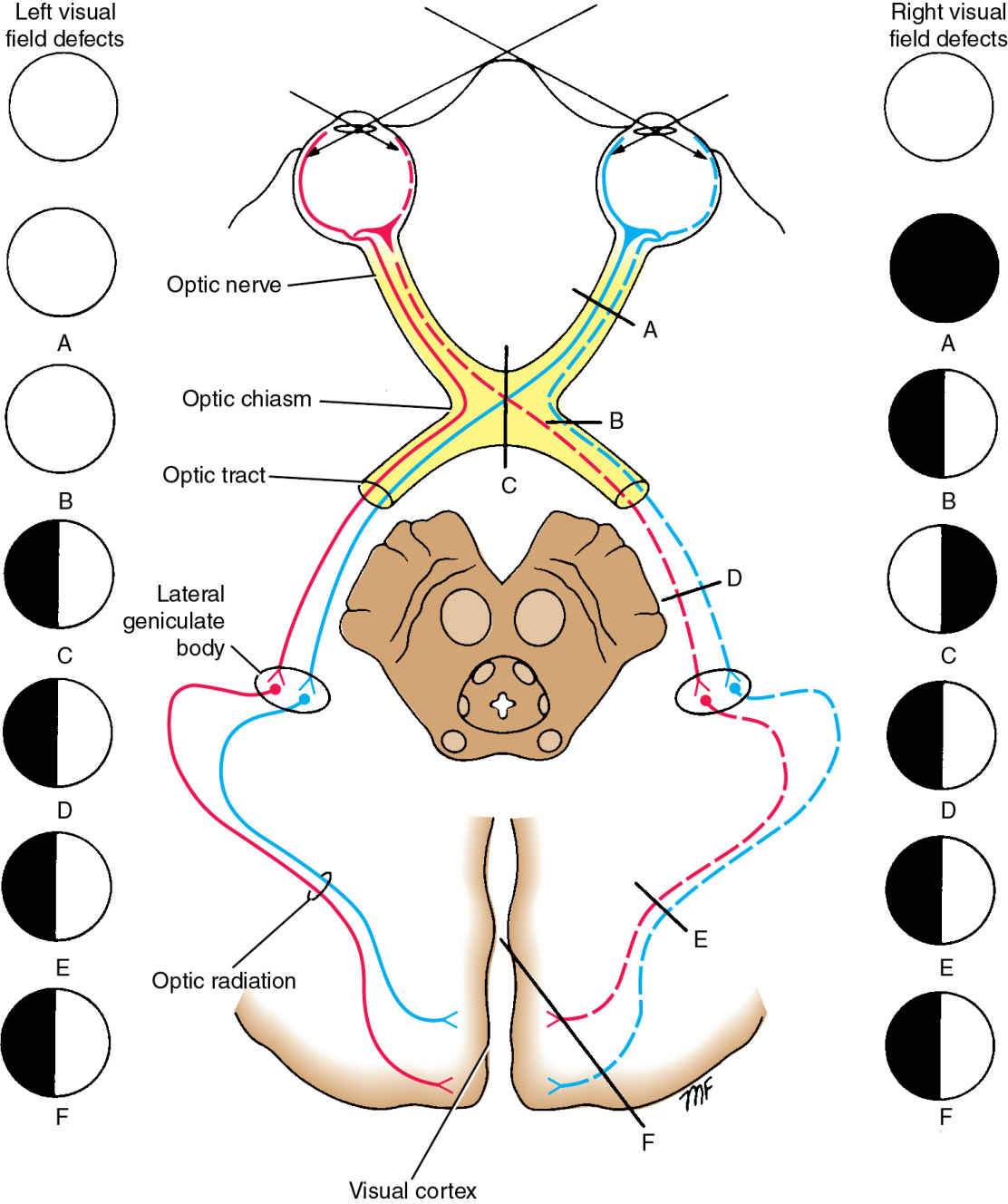
FIG. 19.5 Optic pathway and associated visual field deficits.(A) Lesions of the optic nerves result in total ipsilateral blindness. (B) A lesion at the lateral aspect of the optic chiasm will result in loss of the nasal visual field in the ipsilateral eye. (C) A complete lesion at the optic chiasm centrally causes bitemporal hemianopsia. (D-F) A lesion of the optic tract, optic radiation, or visual cortex will result in loss of the contralateral visual field in both eyes (left homonymous hemianopia in this case).
(Adapted from Snell RS. Clinical Neuroanatomy. 6th ed. Philadelphia, PA: Lippincott Williams & Wilkins; 2006.)
Oculomotor nerve (CN III)
There are two brainstem nuclei that supply the oculomotor nerve: the main motor nucleus (located in the midbrain at the level of the superior colliculus and ventral to the cerebral aqueduct) and the Edinger-Westphal parasympathetic nucleus (situated just ventral and rostral to the main motor nucleus). The nerve fibers from both nuclei travel anteriorly through the medial aspect of the red nucleus and exit the anterior surface of the midbrain along the medial aspect of the cerebral peduncles into the interpeduncular cistern (Fig. 19.6A). The cisternal segments of CN III then pass between the posterior cerebral arteries and the superior cerebellar arteries (Fig. 19.6B), coursing forward inferolateral to the posterior communicating artery to enter the cavernous sinus. After traveling in the lateral wall of the cavernous sinus (Fig. 19.7), CN III then enters the superior orbital fissure and finally the orbit.
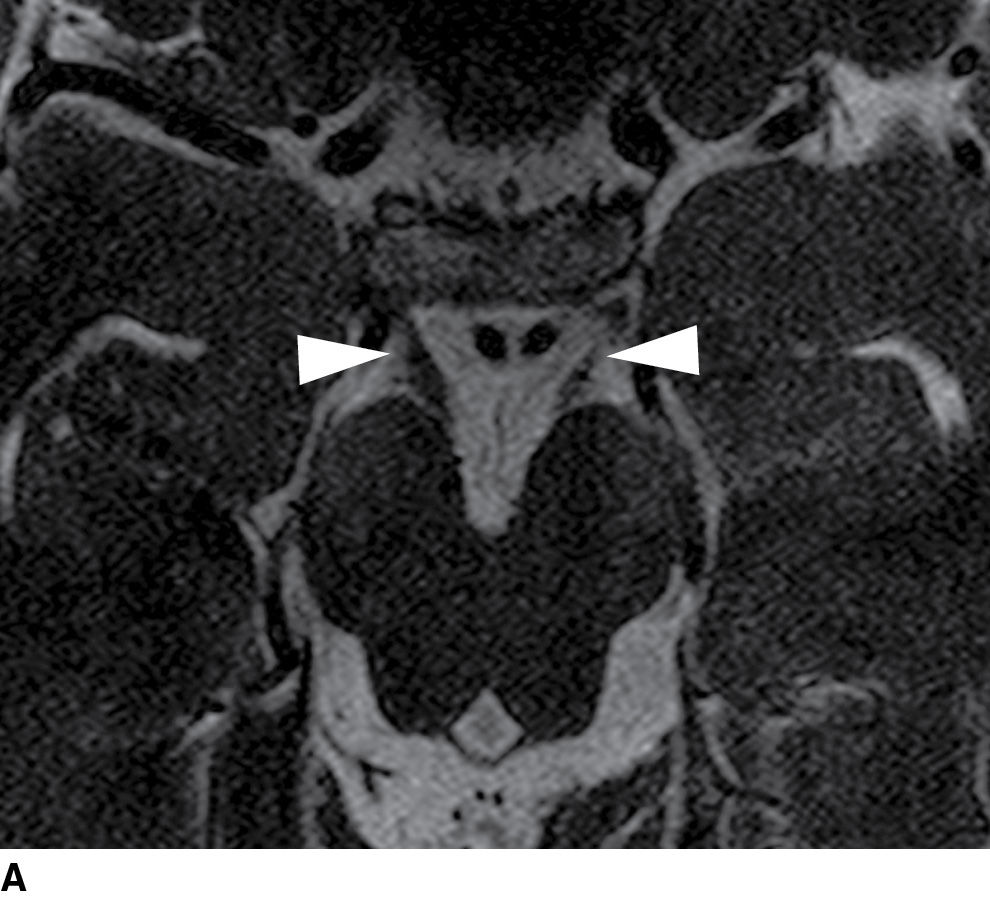
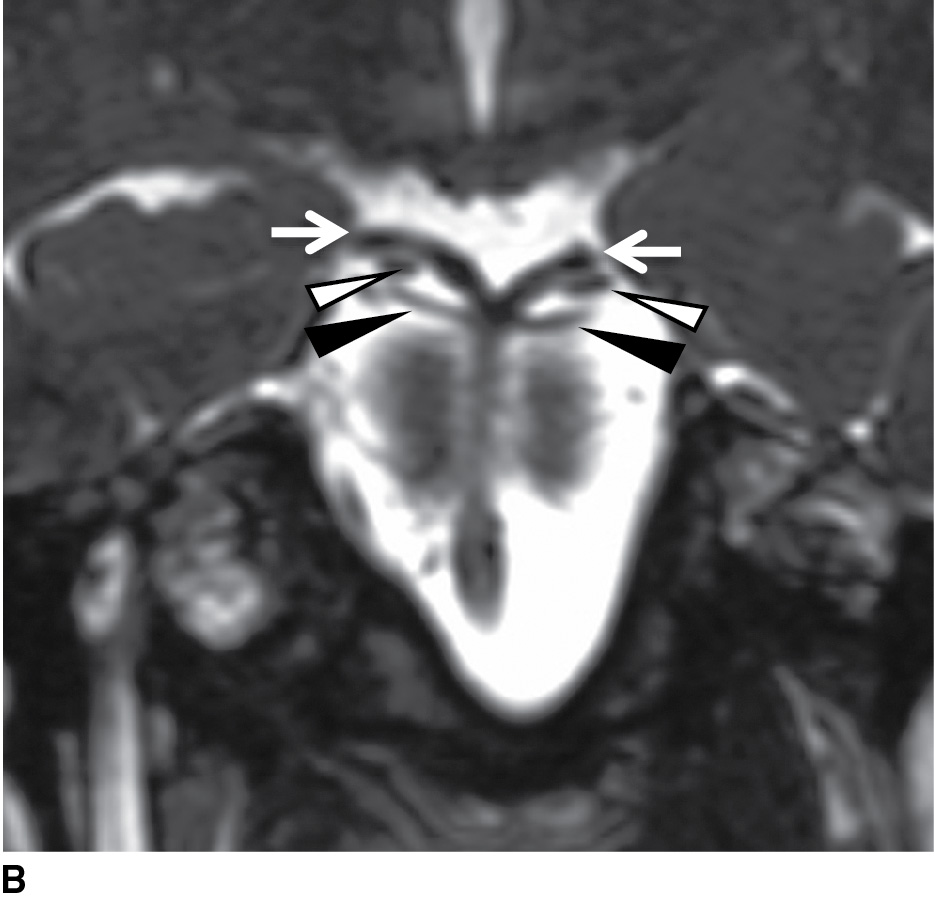
FIG. 19.6 Cisternal segment of the oculomotor nerve. A: Heavily T2-weighted axial image at the level of the midbrain demonstrates the oculomotor nerves (arrowheads) as they exit the interpeduncular fossa and travel anteriorly toward the cavernous sinuses. B: A coronal reformatted heavily T2-weighted image at the level of the top of the basilar artery demonstrates the oculomotor nerves (white arrowheads) traveling between the posterior cerebral arteries superiorly (arrows) and the superior cerebellar arteries inferiorly (black arrowheads).
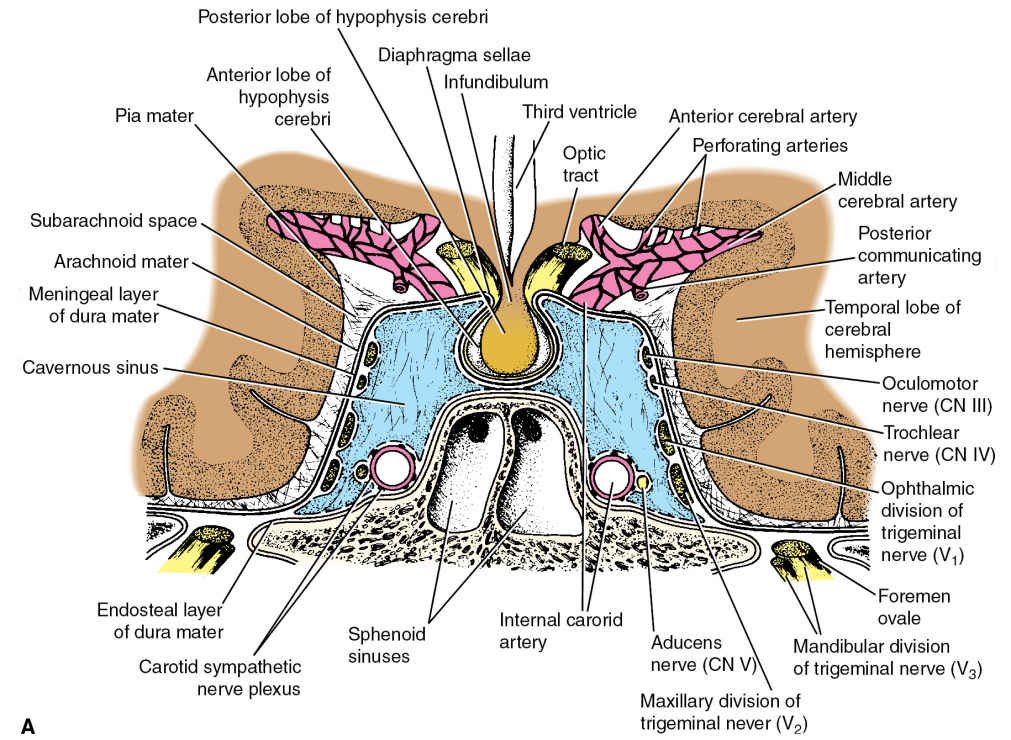
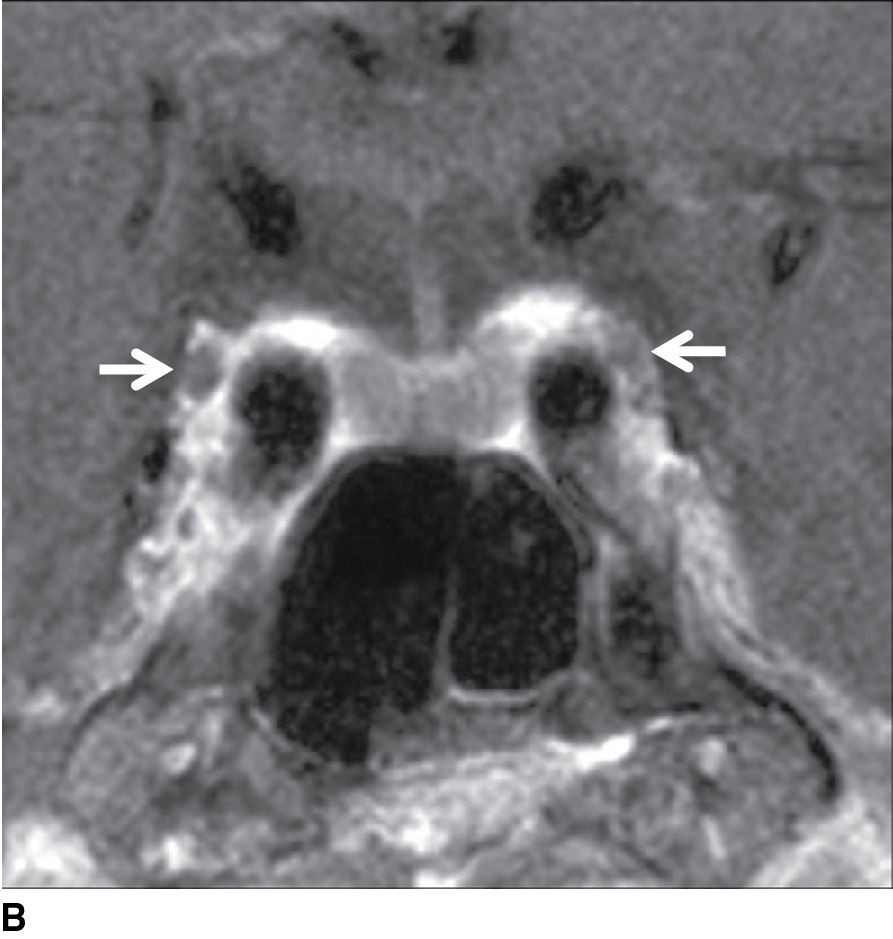
FIG. 19.7 Cavernous sinuses. A: Cranial nerves III, IV, V1 (ophthalmic nerve), and V2 (maxillary nerve) travel in the lateral wall of the cavernous sinus. CN VI lies more medially, within the cavernous sinus itself. B: Coronal T1-weighted image without contrast demonstrates the normal appearance of CN III (arrows) in the cavernous sinus.
(A: From Snell RS. Clinical Neuroanatomy. 6th ed. Philadelphia, PA: Lippincott Williams & Wilkins; 2006.)
Trochlear nerve (CN IV)
The trochlear nucleus is located within the midbrain, caudal to the oculomotor nuclei and ventral to the cerebral aqueduct, at the level of the inferior colliculus. The nerve fibers then pass posteriorly around the periaqueductal gray matter, decussate in the superior medullary velum of the midbrain, and exit at the dorsal surface of the midbrain. Not only does the trochlear nerve have the smallest caliber of any of the cranial nerves, but also it is the only nerve to exit the dorsal surface of the brainstem and has the longest intracranial course. Due to its small size, the cisternal trochlear nerve can be difficult to visualize, even with high-resolution heavily T2-weighted imaging (Fig. 19.8). As with CN III, the trochlear nerve travels in the lateral wall of the cavernous sinus (see Fig. 19.7A) before passing through the superior orbital fissure into the orbit to innervate the superior oblique muscle.
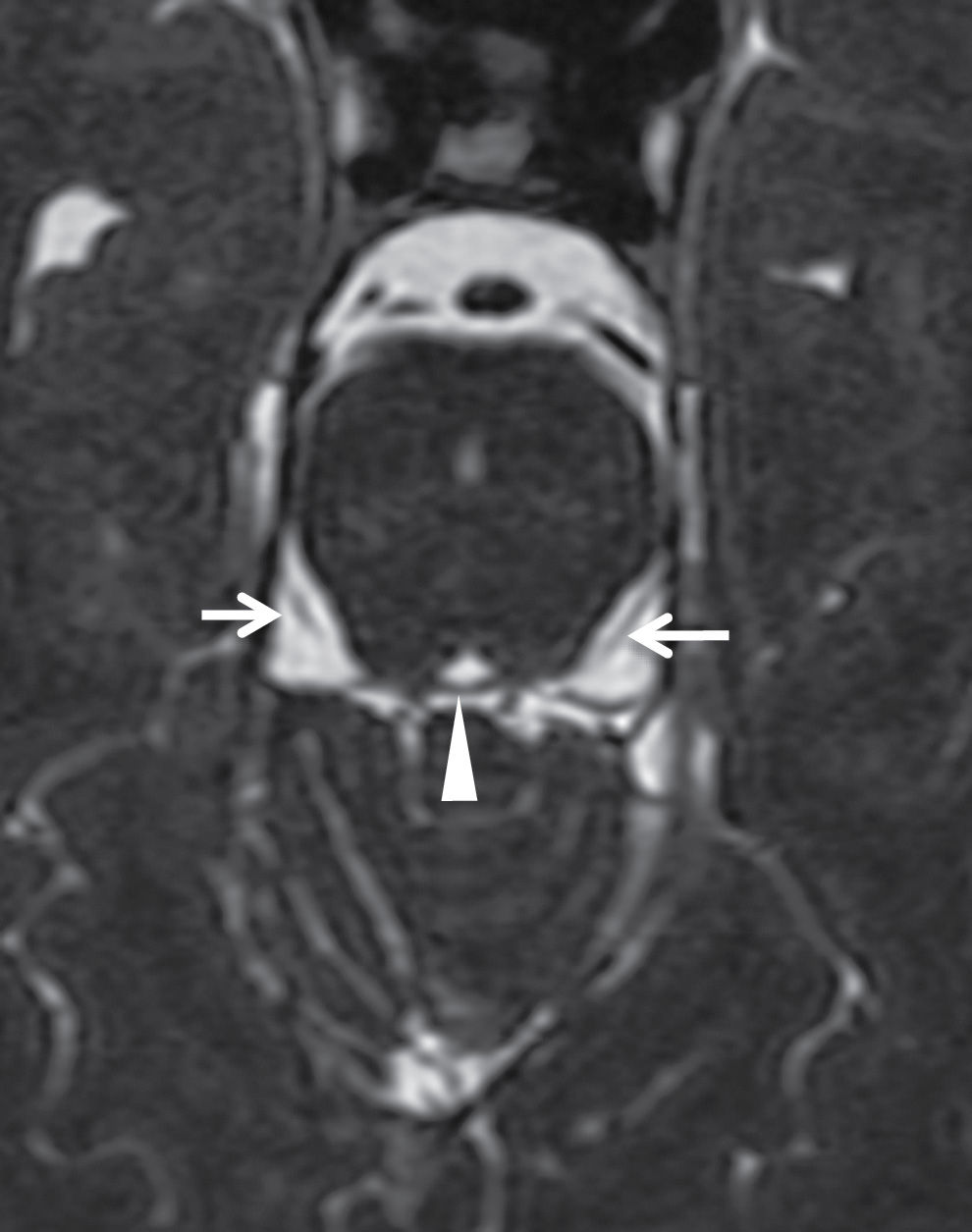
FIG. 19.8 Cisternal trochlear nerve. Heavily T2-weighted axial image at the level of the inferior midbrain demonstrates the superior medullary velum where the decussation of the trochlear nerve fibers occurs (arrowhead) as well as the cisternal segments of the trochlear nerves in the ambient cisterns (arrows).
Trigeminal nerve (CN V)
The principal sensory nucleus of the trigeminal nerve is located within the pons (see Fig. 19.2); however, additional sensory nuclei also extend superiorly and inferiorly within the midbrain (mesencephalic nucleus) and the medulla and upper cervical spinal cord (spinal nucleus). The motor nucleus of the trigeminal nerve is medial and ventral to the principal sensory nucleus in the pons. CN V leaves the ventrolateral aspect of the pons and passes through the cerebellopontine angle cistern to enter Meckel cave, a CSF-filled space just posterior and inferolateral to the cavernous sinus that encloses the ganglion and all three divisions of the trigeminal nerve. The first two divisions of the trigeminal nerve (V1 and V2) are purely sensory and travel in the lateral aspect of the cavernous sinus (see Fig. 19.7A) and exit the skull base through the superior orbital fissure (V1) and foramen rotundum (V2). The mandibular division (V3) is both a motor and sensory nerve and does not enter the cavernous sinus but exits the skull base directly from Meckel cave via foramen ovale. A discussion of the extracranial branches of the trigeminal nerve is beyond the scope of this chapter, but Figure 19.9 provides an overview of the distribution of CN V. The major branch of V1 that is important in imaging is the frontal nerve, which travels in the superior orbit, above the superior rectus/levator palpebrae muscle complex, and is difficult to see normally but can be enlarged in pathologic conditions such as perineural spread of tumor. After exiting foramen rotundum, V2 enters the pterygopalatine fossa (PPF), where it gives off multiple branches to supply sensory innervation to the maxillofacial region below the orbits, including the infraorbital nerve, which continues anteriorly through the inferior orbital fissure and infraorbital foramen. At imaging, the fat in the PPF and size of the infraorbital foramen are important to evaluate when excluding disease of V2 (see Fig. 19.26). At the skull base, the mandibular nerve (V3) lies in the trigeminal fat pad, which is a layer of fat along the medial margin of the lateral pterygoid muscle that is contiguous with foramen ovale and is also an important marker at imaging. Two of the most important V3 branches to evaluate at imaging are the inferior alveolar nerve, which descends next to the mandibular ramus and enters the mandibular canal at the mandibular foramen, and the auriculotemporal nerve, which curves posteriorly around the neck of the mandible and has connections with the facial nerve in the parotid gland (a common site of perineural tumor spread).
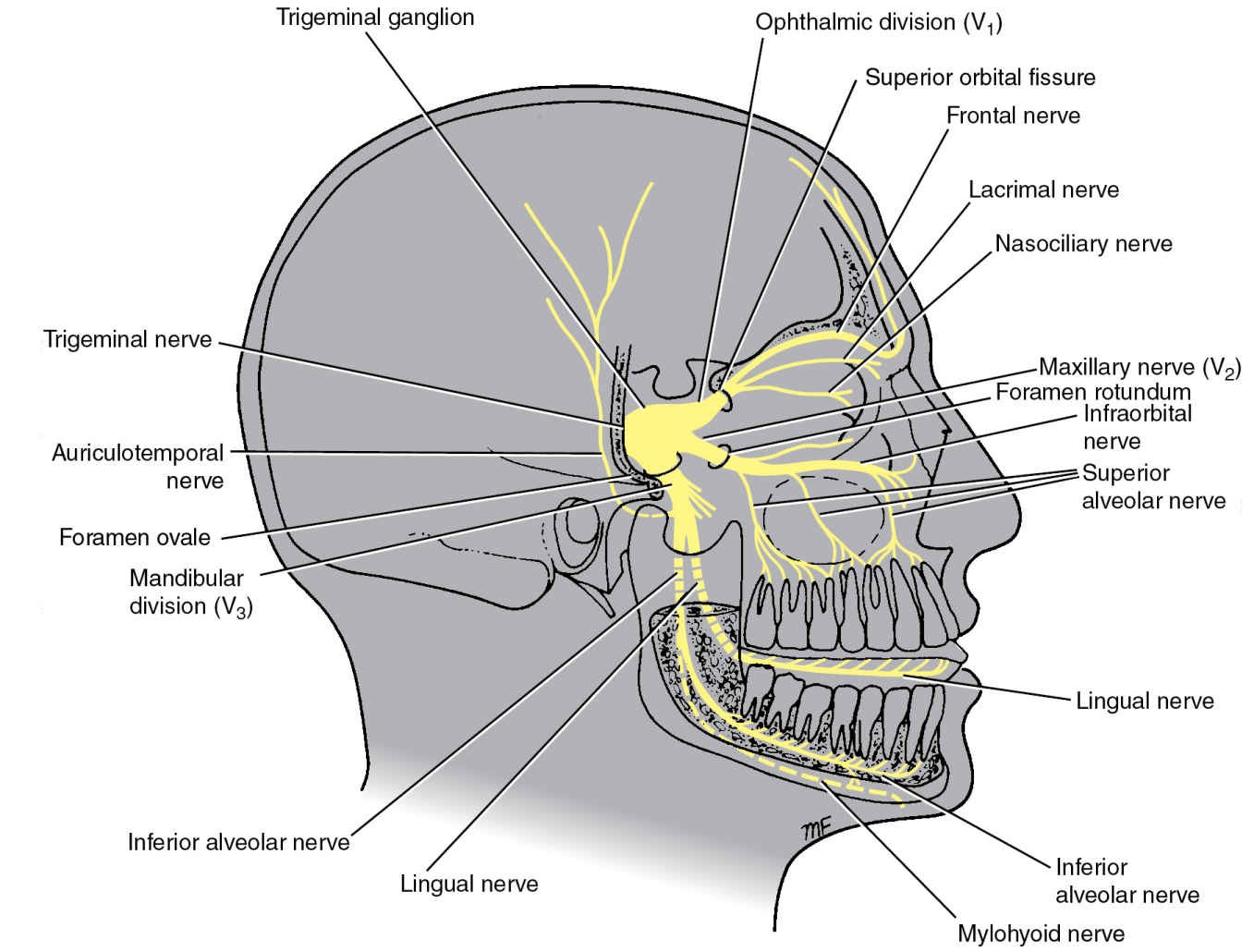
FIG. 19.9 Overview of the trigeminal nerve. (From Snell RS. Clinical Neuroanatomy. 6th ed. Philadelphia, PA: Lippincott Williams & Wilkins; 2006.)
Abducens nerve (CN VI)
The abducens nucleus is a small motor nucleus near the midline in the dorsal pons along the upper aspect of the floor of the fourth ventricle, at the level of the facial colliculus. The nerve exits at the pontomedullary junction just lateral to the pyramids. The cisternal segment of the nerve ascends in the prepontine cistern and enters Dorello canal, a fibro-osseous canal bounded by the petroclinoid ligament superiorly and the petrous apex inferolaterally. CN VI then continues into the cavernous sinus itself (rather than within the dura of the lateral wall as with CN III, IV, V1, and V2), just lateral to the internal carotid artery and medial to V1 (see Fig. 19.7A). The abducens nerve continues through the superior orbital fissure to the orbit to innervate the lateral rectus muscle.
Facial nerve (CN VII)
The motor portion of the facial nerve originates in the caudal pontine tegmentum, just ventrolateral to the CN VI nucleus. The motor fibers loop dorsally around the CN VI nucleus, creating the bump on the floor of the fourth ventricle called the facial colliculus. The parasympathetic (superior salivary) and special sensory (solitary) nuclei lie just dorsolateral to the motor nucleus and contribute fibers to the nervus intermedius. Both the motor branch and the nervus intermedius exit at the anterolateral pontomedullary junction and travel in the cerebellopontine angle cistern to the internal auditory canal. The facial nerve continues through the temporal bone, exiting the skull base at the stylomastoid foramen and entering the parotid gland (see Fig. 19.25). The intratemporal portion of the facial nerve is composed of labyrinthine, geniculate, tympanic, and mastoid segments (Fig. 19.10).
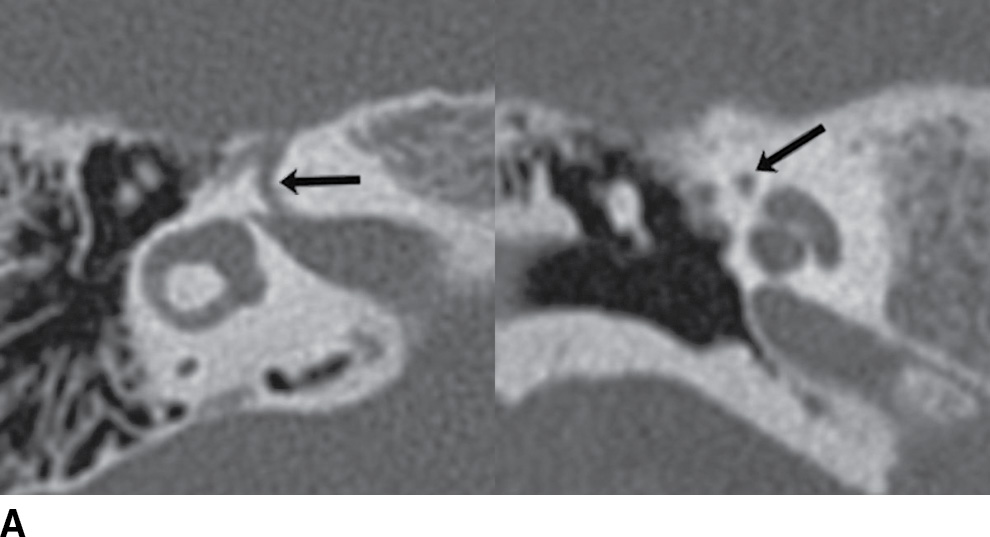
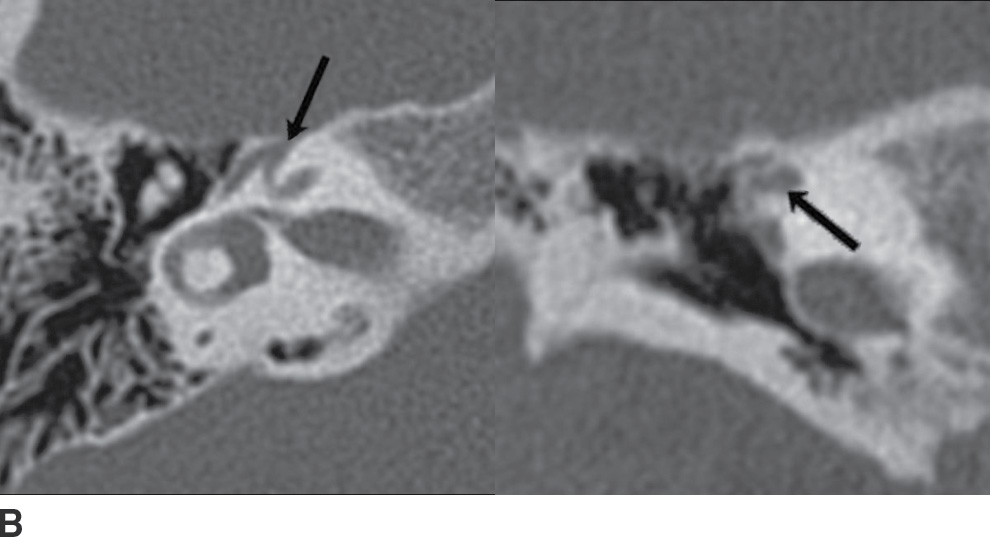
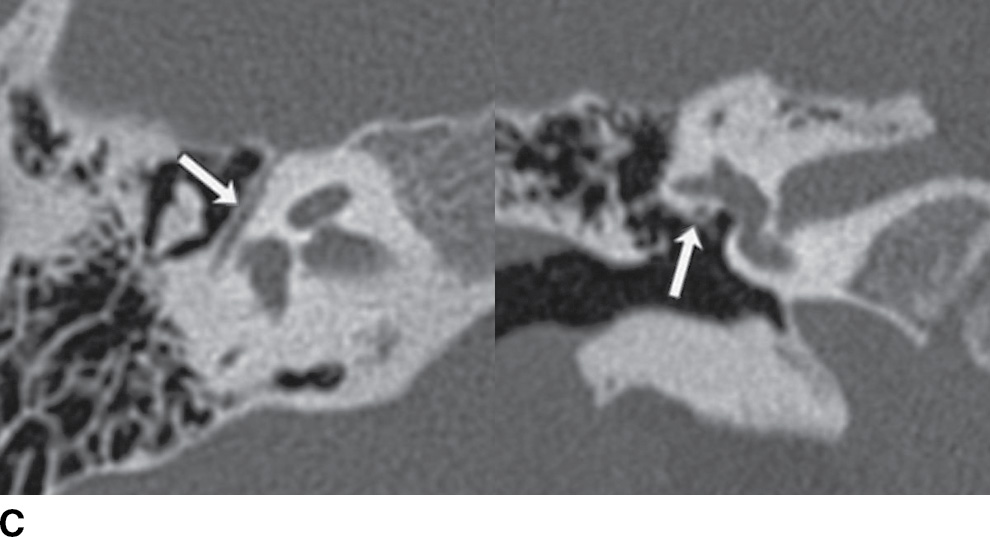
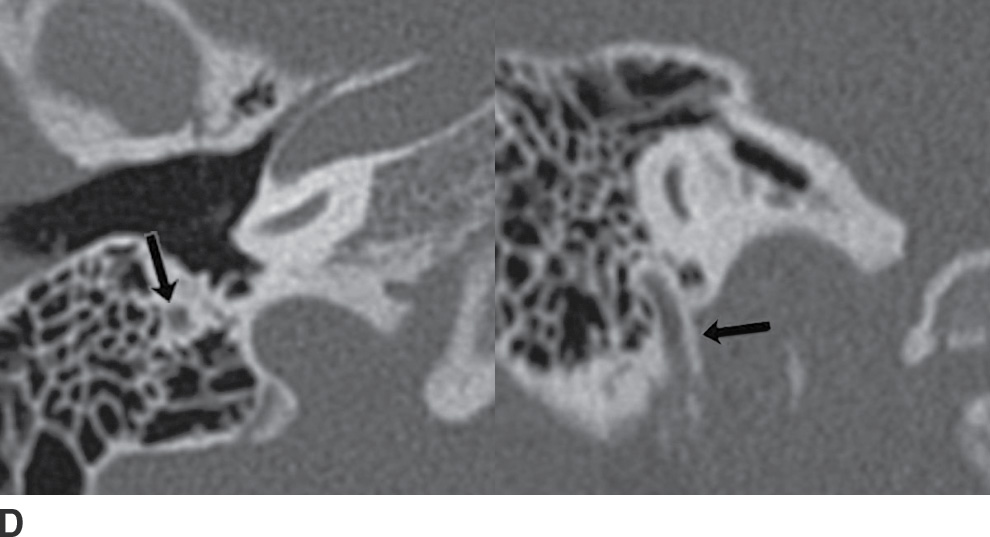
FIG. 19.10 Intratemporal facial nerve. Within the temporal bone, the facial nerve describes an intricate course, with multiple segments and two sharp turns. The “anterior genu” is located at the anterior margins of the temporal bone, at the site of the geniculate ganglion. The “posterior genu” is a 90-degree turn at the posterior margin of the middle ear cavity, where the facial nerve turns inferiorly to course through the mastoid. Arrows in the axial and coronal CT images indicate the normal appearance of the labyrinthine (A), geniculate (B), tympanic (C), and mastoid (D) segments of the facial nerve.
Vestibulocochlear nerve (CN VIII)
Four vestibular nuclei (superior, inferior, medial, and lateral) lie along the floor of the fourth ventricle in the lower pons. More laterally, the two cochlear nuclei (dorsal and ventral) are located at the anterolateral margin of the inferior cerebellar peduncle. The fibers join at the lateral aspect of the pontomedullary junction, traversing the cerebellopontine angle cistern and internal auditory canal with the facial nerve (see Fig. 2A in Chapter 20) to reach the inner ear structures. In the internal auditory canal, the facial nerve is in the anterior superior quadrant, the cochlear nerve is in the anterior inferior quadrant, and the superior and inferior vestibular nerves travel posteriorly (Fig. 19.11).
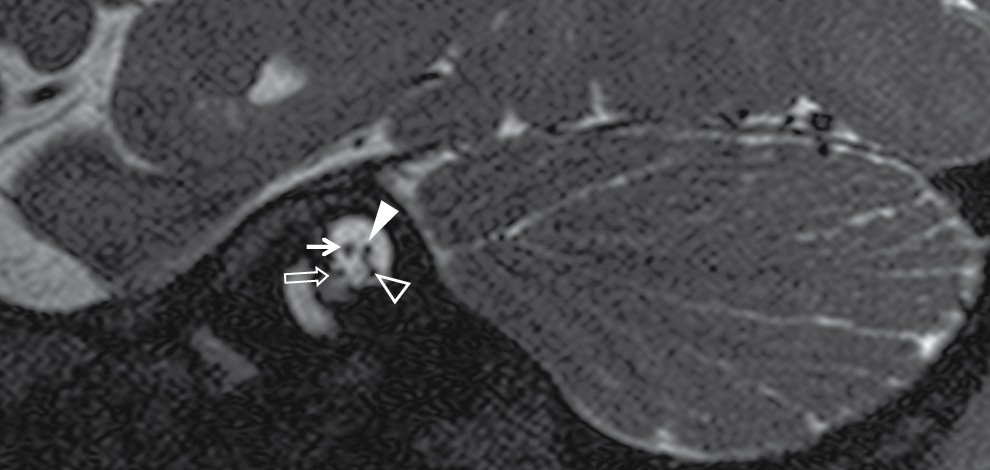
FIG. 19.11 CN VII and VIII in the internal auditory canal. In the internal auditory canal, the facial nerve (thin arrow) is in the anterior superior quadrant, the cochlear nerve (open arrow) is in the anterior inferior quadrant (just entering the cochlea in this image), and the superior (solid arrowhead) and inferior (open arrowhead) vestibular nerves travel posteriorly (seen as a confluent single nerve posteriorly here, separating more clearly at the far lateral aspect of the IAC).
Glossopharyngeal nerve (CN IX)
Fibers from three nuclei in the medulla contribute to the glossopharyngeal nerve: the motor nucleus ambiguus (also shared with X and XI), the solitary tract nucleus (taste, shared with VII and X), and the inferior salivary nucleus (parasympathetic innervation to the parotid). Sensory fibers from the pharynx also travel through CN IX but synapse in the spinal nucleus of the trigeminal nerve. The glossopharyngeal nerve exits the medulla as a collection of rootlets in a groove between the olive and the inferior cerebellar peduncle (the postolivary sulcus) and then exits the skull base with CN X and XI via the jugular foramen. Just before exiting the skull base, CN IX gives off a branch to the middle ear, the tympanic branch or Jacobson nerve, which is responsible for glomus tumors (see Figs. 19.22 and 19.24). Inferior to the skull base, CN IX, X, and XI are situated between the internal carotid artery and internal jugular vein, posterior to the styloid process (in the poststyloid parapharyngeal space, or carotid space; see Fig. 19.23).
Vagus nerve (CN X)
The vagus nerve has three nuclei in the medulla: a main motor nucleus (nucleus ambiguus, shared with IX and XI), a parasympathetic nucleus (dorsal motor nucleus of CN X), and a sensory nucleus (solitary tract nucleus, responsible for taste and shared with CN VII and IX). Similar to CN IX, the vagus nerve exits the medulla in the postolivary sulcus and exits the skull through the jugular foramen. An auricular branch (Arnold nerve) ascends from the jugular foramen to the temporal bone and can also give rise to glomus tumors (Figs. 19.22 and 19.24). The vagus nerve has the longest course of the cranial nerves and has the most extensive distribution, initially traveling in the carotid sheath between the internal and common carotid arteries and internal jugular vein before carrying parasympathetic and visceral sensory fibers to the thorax and abdomen. Important branches to the larynx are the recurrent laryngeal nerves, which loop under the aortic arch on the left and under the subclavian artery on the right.
Spinal accessory nerve (CN XI)
CN XI is a motor nerve that is often described as having a cranial root and a spinal root. The nucleus ambiguus (shared with IX and X) supplies the cranial root, which travels separately from the spinal root in the jugular foramen and ultimately joins the vagus nerve. The spinal root is supplied by a nucleus in the ventral horn of the upper five segments of the cervical spinal cord. The spinal rootlets exit the cervical cord to form a single root that ascends through the foramen magnum to then exit the skull base via the jugular foramen and travel in the carotid sheath before supplying the sternocleidomastoid and trapezius muscles.
Jugular foramen
The jugular foramen is most often described as having two compartments: an anteromedial pars nervosa and a posterolateral pars vascularis, partially separated by bony spines on opposite surfaces of the temporal and occipital bones, the intrajugular processes. Cranial nerve IX and its tympanic branch (Jacobson nerve) as well as the inferior petrosal sinus typically travel in the medial aspect (pars nervosa), while cranial nerve X, its auricular branch (Arnold nerve), cranial nerve XI, and the internal jugular vein travel in the lateral aspect (pars vascularis). However, some prefer not to use these terms as the anatomy can vary, and in some cases, both cranial nerves IX and X may travel in the medial aspect. For simplicity, the nerves and the inferior petrosal sinus can be thought of as traveling medially, while the internal jugular vein always travels laterally.
Hypoglossal nerve (CN XII)
The hypoglossal nucleus is a somatic motor nucleus located close to the midline in the medulla, immediately ventral to the floor of the inferior fourth ventricle. The nerve fibers then run anteriorly to exit the medulla in the preolivary sulcus (between the pyramid and the olive), cross the posterior cranial fossa anteriorly, and exit the skull base through the hypoglossal canal. In the neck, the nerve descends between the internal carotid artery and internal jugular vein, medial to CN IX, X, and XI, and then turns anteriorly at the posterior belly of the digastric muscle and passes deep to the posterior margin of the mylohyoid muscle, supplying the majority of the extrinsic muscles and all of the intrinsic muscles of the tongue.
Imaging Protocols
Magnetic resonance imaging is the study of choice for imaging the cranial nerves (1–3). However, CT can be complimentary when evaluating the bony margins of lesions at the skull base (2). CT is also necessary when evaluating a distal vagal neuropathy as a potential cause for vocal cord paralysis as imaging must extend to the right subclavian artery (right recurrent laryngeal nerve) and/or AP window (left recurrent laryngeal nerve).
When designing a protocol to evaluate a specific cranial nerve, it is crucial to cover the entire anatomic extent with careful attention to its location in the brainstem, its cisternal course, its associated foramen/fissure and adjacent fat pads at the skull base, as well as its extracranial course. Multiplanar high-resolution MR imaging is mandatory. High-resolution heavily T2-weighted 3D sequences (e.g., CISS, FIESTA, DRIVE) are customary for examination of the cisternal segments (1,2,4). Table 19.3 summarizes the additional imaging considerations for covering the extracranial portions of the various cranial nerves. Gadolinium is used to evaluate for inflammation, infection, and/or leptomeningeal disease. The cranial nerves do not normally enhance (with the exception of portions of CN VII) and should be isointense to the brain parenchyma. MR angiography may be added if an aneurysm is suspected.
Table 19.3 Imaging the Extracranial Course of the Cranial Nerves
Pathology
Knowledge of the anatomy in the context of the clinical exam and functional deficits in an individual patient are the most important assets to the radiologist evaluating the cranial nerves. Localization of the abnormality, in combination with the specific CT and MR findings, can usually narrow the differential diagnosis to a very short list of possibilities. The major pathologic entities that affect the cranial nerves are discussed below; however, Table 19.4 provides a more comprehensive list of the diseases that can affect their various anatomic segments.
Table 19.4 Differential diagnosis of cranial nerve pathology by anatomic location

Neoplasms
Esthesioneuroblastoma
Esthesioneuroblastoma (or olfactory neuroblastoma) is a rare malignant neuroectodermal tumor arising from the olfactory mucosa in the superior nasal cavity (5,6). There is a bimodal age distribution, with peaks in the second and fifth decades (7). Clinical presentation typically includes nasal congestion, epistaxis, anosmia, and headache. On CT, these lesions are hyperdense and will enhance homogeneously, with or without adjacent bony destruction, especially of the cribriform plate. The classic imaging description is of a “dumbbell-shaped” mass centered at the cribriform plate, with extension into the anterior cranial fossa and superior nasal cavity. They are intermediate in signal intensity on both T1- and T2-weighted images with intense homogeneous enhancement. Peripheral cysts at the margins of the intracranial component of the tumor have been described as a characteristic MR finding (8) (Fig. 19.12).
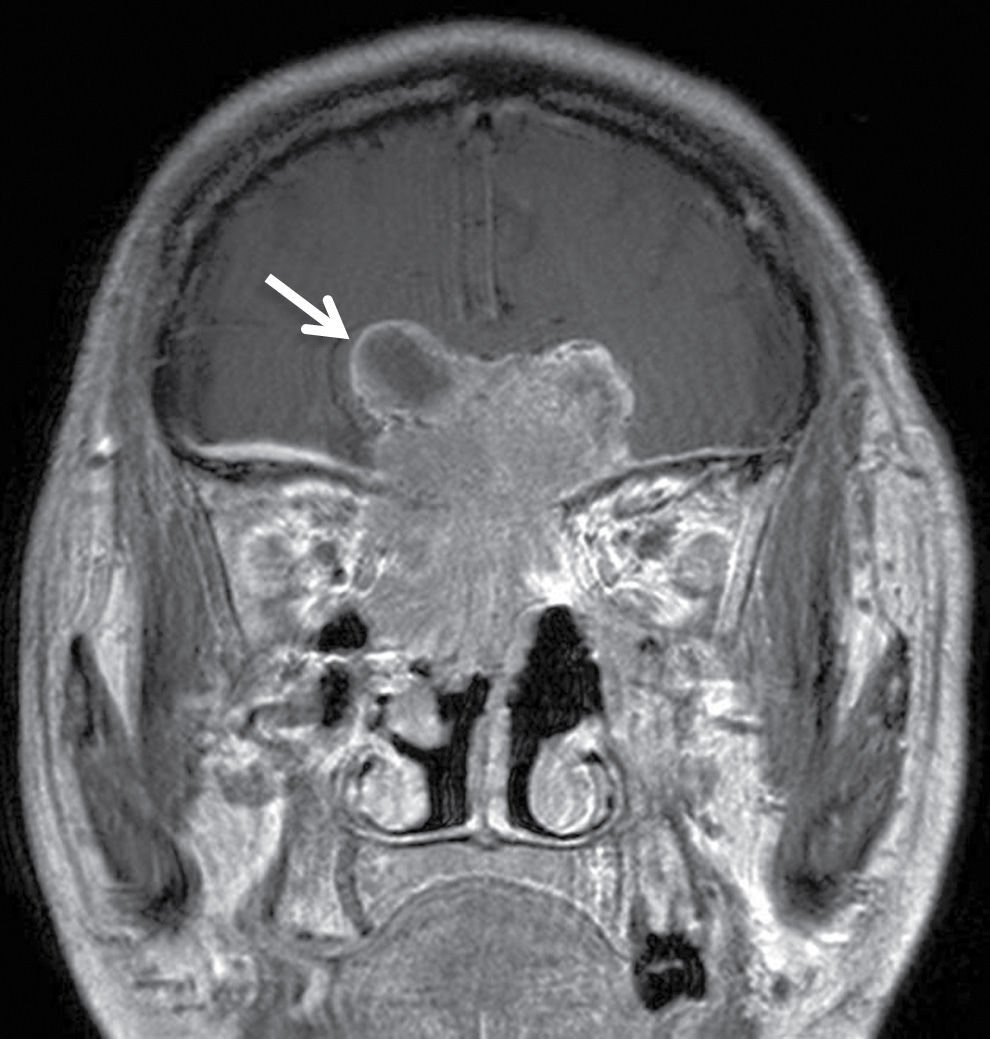
FIG. 19.12 Esthesioneuroblastoma. Coronal T1-weighted postcontrast image demonstrates an avidly enhancing mass lesion centered at the cribriform plate with involvement of the anterior cranial fossa, superior nasal cavity, and right orbit. A characteristic peripheral cystic component is seen along the right superolateral aspect of the lesion intracranially (arrow).
Related posts:
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


